
eBook - ePub
Practical Approaches to Biological Inorganic Chemistry
- 336 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Practical Approaches to Biological Inorganic Chemistry
About this book
The book reviews the use of spectroscopic and related methods to investigate the complex structures and mechanisms of biological inorganic systems that contain metals. Each chapter presents an overview of the technique including relevant theory, clearly explains what it is and how it works and then presents how the technique is actually used to evaluate biological structures. Practical examples and problems are included to illustrate each technique and to aid understanding. Designed for students and researchers who want to learn both the basics, and more advanced aspects of bioinorganic chemistry.
- Many colour illustrations enable easier visualization of molecular mechanisms and structures
- Worked examples and problems are included to illustrate and test the reader's understanding of each technique
- Written by a multi-author team who use and teach the most important techniques used today to analyse complex biological structures
Information
Topic
Physical SciencesSubtopic
Inorganic ChemistryChapter 1
An Overview of the Roles of Metals in Biological Systems
Robert R. Crichton, Batiment Lavoisier, Université Catholique de Louvain, Louvain-la-Neuve, Belgium
Chapter Outline
Introduction: Which Metals Ions and Why?
Some Physicochemical Considerations on Alkali Metals
Na+ and K+ – Functional Ionic Gradients
Mg2+ – Phosphate Metabolism
Ca2+ and Cell Signalling
Zinc – Lewis Acid and Gene Regulator
Iron and Copper – Dealing with Oxygen
Ni and Co – Evolutionary Relics
Mn – Water Splitting and Oxygen Generation
Mo and V – Nitrogen Fixation
Introduction: Which Metals Ions and Why?
In the companion book to this one, ‘Biological Inorganic Chemistry 2nd edition’ (Crichton, 2011), we explain in greater detail why life as we know it would not be possible with just the elements found in organic chemistry – namely carbon, oxygen, hydrogen, nitrogen, phosphorus and sulfur. We also need components of inorganic chemistry as well, and in the course of evolution nature has selected a number of metal ions to construct living organisms. Some of them, like sodium and potassium, calcium and magnesium, are present at quite large concentrations, constituting the so-called ‘bulk elements’, whereas others, like cobalt, copper, iron and zinc, are known as ‘trace elements’, with dietary requirements that are much lower than the bulk elements.
Just six elements – oxygen, carbon, hydrogen, nitrogen, calcium and phosphorus – make up almost 98.5% of the elemental composition of the human body by weight. And just 11 elements account for 99.9% of the human body (the five others are potassium, sulfur, sodium, magnesium and chlorine). However, between 22 and 30 elements are required by some, if not all, living organisms, and of these are quite a number are metals. In addition to the four metal ions mentioned above, we know that cobalt, copper, iron, manganese, molybdenum, nickel, vanadium and zinc are essential for humans, while tungsten replaces molybdenum in some bacteria. The essential nature of chromium for humans remains enigmatic.
Just why these elements out of the entire periodic table (Figure 1.1) have been selected will be discussed here. However, their selection was presumably based not only on suitability for the functions that they are called upon to play in what is predominantly an aqueous environment, but also on their abundance and their availability in the earth’s crust and its oceans (which constitute the major proportion of the earth’s surface).
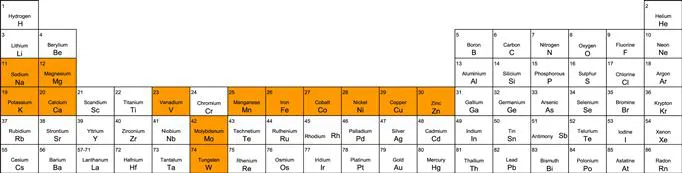
FIGURE 1.1 An abbreviated periodic table of the elements showing the metal ions discussed in this chapter.
The 13 metal ions that we will discuss here fall naturally into four groups based on their chemical properties. In the first, we have the alkali metal ions Na+ and K+. Together with H+ and Cl−, they bind weakly to organic ligands, have high mobility, and are therefore ideally suited for generating ionic gradients across membranes and for maintaining osmotic balance. In most mammalian cells, most K+ is intracellular, and Na+ extracellular, with this concentration differential ensuring cellular osmotic balance, signal transduction and neurotransmission. Na+ and K+ fluxes play a crucial role in the transmission of nervous impulses both within the brain and from the brain to other parts of the body.
The second group is made up by the alkaline earths, Mg2+ and Ca2+. With intermediate binding strengths to organic ligands, they are, at best semi-mobile, and play important structural roles. The role of Mg2+ is intimately associated with phosphate, and it is involved in many phosphoryl transfer reactions. Mg-ATP is important in muscle contraction, and also functions in the stabilisation of nucleic acid structures, as well as in the catalytic activity of ribozymes (catalytic RNA molecules). Mg2+ is also found in photosynthetic organisms as the metal centre in the light-absorbing chlorophylls. Ca+ is a crucial second messenger, signalling key changes in cellular metabolism, but is also important in muscle activation, in the activation of many proteases, both intra- and extracellular, and as a major component of a range of bio-minerals, including bone.
Zn2+, which is arguably not a transition element,1 constitutes the third group on its own. It is moderate to strong binding, is of intermediate mobility and is often found playing a structural role, although it can also fulfil a very important function as a Lewis acid. Structural elements, called zinc fingers, play an important role in the regulation of gene expression.
The other eight transition metal ions, Co, Cu, Fe, Mn, Mo, Ni, V and W form the final group. They bind tightly to organic ligands and therefore have very low mobility. Since they can exist in various oxidation states, they participate in innumerable redox reactions, and many of them are involved in oxygen chemistry. Fe and Cu are constituents of a large number of proteins involved in electron transfer chains. They also play an important role in oxygen-binding proteins involved in oxygen activation as well as in oxygen transport and storage. Co, together with another essential transition metal, Ni, is particularly important in the metabolism of small molecules like carbon monoxide, hydrogen and methane. Co is also involved in isomerisation and methyl transfer reactions. A major role of Mn is in the catalytic cluster involved in the photosynthetic oxidation of water to dioxygen in plants, and, from a much earlier period in geological time, in cyanobacteria. Mo and W enzymes contain a pyranopterindithiolate cofactor, while nitrogenase, the key enzyme of N2 fixation contains a molybdenum–iron–sulfur cofactor, in which V can replace Mo when Mo is deficient. Other V enzymes include haloperoxidases. To date no Cr-binding proteins have been found, adding to the lack of biochemical evidence for a biological role of the enigmatic Cr.
Some Physicochemical Considerations on Alkali Metals
Before considering, in more detail, the roles of the alkali metals, Na+ and K+, and the alkaline earth metals, Mg2+ and Ca2+, it may be useful to examine some of their physicochemical properties (Table 1.1). We can observe, for example that Na+ and K+ have quite significantly different unhydrated ionic radii, whereas, the hydrated radii are much more similar. It therefore comes as no surprise that the pumps and channels which carry them across membranes, and which can easily distinguish between them, as we will see shortly, transport the unhydrated ions. Although not indicated in the table, it is clear that Na+ is invariably hexa-coordinate, whereas K+ and Ca2+ can adjust to accommodate 6, 7 or 8 ligands. As we indicated above, both Na+ and K+ are characterised by very high solvent exchange rates (around 109/s), consistent with their high mobility and their role in generating ionic gradients across membranes. In contrast, the mobility of Mg2+ is some four orders of magnitude slower, consistent with its essentially structural and catalytic. Perhaps surprisingly, Ca2+ has a much higher mobility (3 × 108/s), which explains why it is involved in cell signalling via rapid changes on Ca2+ fluxes.
TABLE 1.1
Properties of Common Biological Cations
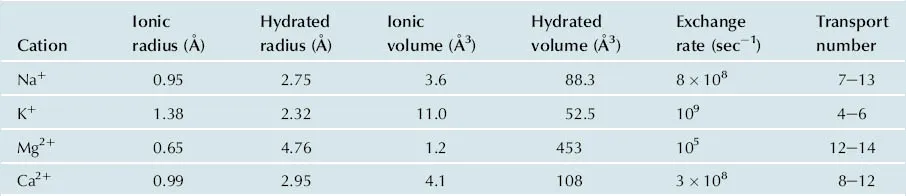
(From Maguire and Cowan, 2002).
The selective binding of Ca2+ by biological ligands compared to Mg2+ can be explained by the difference in their ionic radius, as we pointed out above. Also, for the smaller Mg2+ ion, the central field of the cation dominates its coordination sphere, whereas for Ca2+, the second and possibly even the third, coordination spheres have an important influence resulting in irregular coordination geometry. This allows Ca2+, unlike Mg2+ to bind to a large number of centres at once.
The high charge density on Mg2+ as a consequence of its small ionic radius ensures that it is an excellent Lewis acid in reactions notably involving phosphoryl transfers and hydrolysis of phosphoesters. Typically, Mg2+ functions as a Lewis acid, either by activating a bound nucleophile to a more reactive anionic form (e.g. water to hydroxide anion), or by stabilising an intermediate. The invariably hexacoordinate Mg2+ often participates in structures where the metal is bound to four or five ligands from the protein and a phosphorylated substrate. This leaves one or two coordination positions vacant for occupation by water molecules, which can be positioned in a particular geometry by the Mg2+ to participate in the catalytic mechanism of the enzyme.
Na+ and K+ – Functional Ionic Gradients
How, we might ask, do the pumps and channels responsible for transport across membranes distinguish between Na+ and K+ ions? Studies over the last 50 years or so of synthetic and naturally occurring small molecules which bind ions have established the basic rules of ion selectivity. Two major factors appear to be of capital importance, namely the molecular composition and the stereochemistry (essentially the size) of the binding site. Synthetic molecules have been created which selectivity bind Li+ (radius 0.60 Å), Na+ (0.95 Å), K+ (1.35 ...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Preface
- Chapter 1. An Overview of the Roles of Metals in Biological Systems
- Chapter 2. Introduction to Ligand Field Theory
- Chapter 3. EPR Spectroscopy
- Chapter 4. Introduction to Biomolecular NMR and Metals
- Chapter 5. 57Fe-Mössbauer Spectroscopy and Basic Interpretation of Mössbauer Parameters
- Chapter 6. X-ray Absorption Spectroscopy in Biology (BioXAS)
- Chapter 7. Other Spectroscopic Methods for Probing Metal Centres in Biological Systems
- Chapter 8. An Introduction to Electrochemical Methods for the Functional Analysis of Metalloproteins
- Chapter 9. X-ray Crystallography
- Chapter 10. Genetic and Molecular Biological Approaches for the Study of Metals in Biology
- Index
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Practical Approaches to Biological Inorganic Chemistry by Robert R. Crichton,Ricardo O. Louro in PDF and/or ePUB format, as well as other popular books in Physical Sciences & Inorganic Chemistry. We have over 1.5 million books available in our catalogue for you to explore.