
- 684 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Auditory Physiology and Perception documents the proceedings of the 9th International Symposium on Hearing held in Careens, France, 9-14 June 1991. The aim of the symposium was to promote exchanges between hearing scientists working with different approaches from cell biology to psychology. The volume is organized into 10 parts. Part I contains papers on the biology of inner ear cells. Part II presents studies on auditory periphery functioning. Part III examines frequency selectivity while Part IV contains papers that deal with the subject of pitch. The papers in Part V examine the coding of intensity. Parts VI and VII discuss temporal analyses and spectral shape analysis, respectively. Part VIII takes up spectro-temporal processing. Part IX covers binaural interactions and sound localization. The studies in Part X focus on pathologies, such as the relations between evoked otoacoustic emissions and pure tone audiometry and the effect of short duration acoustic trauma on activity of single neurons in the ventral cochlear nucleus. The final chapter of the text is a tribute to Professor Zwicker, a leading scientist in hearing, who passed away some months before the symposium.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
MedicineSubtopic
PhysiologyAuditory Periphery Functioning
Cochlear Mechanics
J.P. Wilson, Department of Communication & Neuroscience, Keele University, Staffs ST5 5BG, UK
ABSTRACT
From an early time frequency analysis has been thought to be mechanical in origin. The first direct measurements, however, showed consistently poor tuning. This led to the concept of a second filter mechanism beyond the basilar membrane. Electrical tuning properties in some species appeared to support this view. With the discovery of otoacoustic emissions, an earlier hypothesis that mechanical amplification might enhance tuning received support. The most recent mechanical measurements demonstrate the extreme vulnerability of basilar membrane tuning and show that under good conditions it can indeed be adequately sharp. Although the outer hair cells are the prime candidates for the cochlear motor, their mode of action is not yet fully worked out. Furthermore there still appears to be room for other mechanisms in some non-mammalians. Some important aspects of psychophysical phenomena such as masking curves, equal loudness curves, recruitment, hearing loss, distortion products, and pich shifts now appear to be explicable at the mechanical level.
KEYWORDS
Basilar membrane
cochlea
cochlear tuning
frequency analysis
comparative studies
travelling waves
mechanical amplifier
active processes
motility
distortion products
hearing loss
INTRODUCTION
In addition to its obvious function as that of sensory transducer converting the acoustical input into a code of nerve impulses, the cochlea also performs the first stage of sensory analysis separating out the different regions of the frequency spectrum into different channels for parallel transmission to the brain. This “place” analysis is based on mechanical processes but in a way which turns out to be more complex than originally thought, and is still not completely understood in detail. For this reason it is probably helpful to approach the topic from an historical viewpoint explaining how our present understanding has unfolded. This may also be appropriate because in certain respects our present conceptions are closer to the starting point than to some of the points en route.
Inevitably the topic of cochlear mechanics focuses on “place” aspects of hearing rather than temporal aspects. This does not imply that temporal processing is unimportant but rather that temporal features of the signal have to be “seen” through the filters of the cochlea. In certain respects these filters appear to have evolved in such a way that loss of temporal information is minimised.
With parallel evolution of hearing organs it is inevitable that differing mechanisms receive different emphasis in different species and comparative studies may allow a mechanism that is embryonic or vestigial in one species to be understood more readily when observed in a more fully developed form. Where, however, completely different paths are taken, this approach may not be useful.
This review will start with mammalian hearing and consider other developments where these might be appropriate.
There are several areas of research which are closely linked with the present topic which will be discussed in passing, but which are now too large to be considered fully in a review of this length. These include models of the cochlea (see de Boer, 1980, 1984), otoacoustic emissions (see Kemp, 1978, Wilson, 1984), micromechanical processes, and cochlear potentials (see Dallos, this volume).
MAMMALIAN COCHLEAR MECHANICS
History
It is appropriate to start with Hermann von Helmholtz (1863, 1877) because he brought together anatomical findings, a physical model, and psychophysical considerations in a form which was to hold sway for many decades. So persuasive infact, that it probably stifled further research in this area! Helmholtz considered that the basilar membrane consisted of a series of simple second-order resonators each tuned to a slightly different frequency and, invoking Müller’s theory of specific nerve energies, each connected by a separate receptor and nerve fibre to the brain. In 1863 he postulated that the mechanical resonators were the rods of Corti but by 1877 it was realised that birds did not have these and that the structure showing most change in dimension from base to apex of the cochlea was the width of the basilar membrane. Furthermore, the basilar membrane (BM) was observed to split easily along the direction of its transverse fibres. From this, Helmholtz concluded that it could be stretched strongly across its width and very weakly along its length and thereby providing a graded series of independent vibrating strings in which the tuning frequencies were lowered to the appropriate range by the masses of the (mechanically independent) overlying structures. He estimated the amount of mechanical damping in the resonators from psychophysical observations. By considering the lowest tone for which the ear could follow a rapid alternation of notes, he concluded that its Q- factor must be about 25 (Q10dB = 8.4), a result remarkably close to modern estimates! He commented that the illustration (Fig. 1a) that he had derived for intensity as a function of frequency might, with suitable scaling, also represent intensity as a function of position. It is interesting to note that Helmholtz considered his theory partly proved by an observation of V. Hensen that different auditory hairs of the tail of the opossum shrimp were tuned to different frequencies. Hensen used a keyed bugle as acoustic stimulator and perhaps this rates as the first direct observation of mechanical auditory tuning!
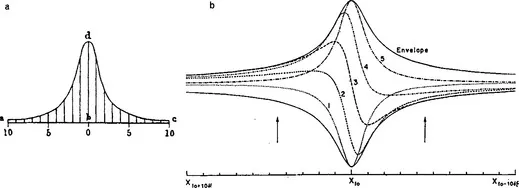
Fig. 1 (a) Helmholtz (1877) resonance curve showing variation of intensity (amplitude)2 over a range (a-c) of four semitones or over a distance of 1.5mm on the BM. (b) Computed instantaneous “travelling wave” patterns (1-5) and envelope (solid) for a bank of independent resonators. The vertical arrows represent adjacent reeds observed by Békésy (1960, Fig. 12.11(a)) which misses the intermediate region of interest.
The principle of the telephone, which had been recently invented, appeared so relevant to mechanisms of hearing that Rutherford (1886) applied it in a theory in which it was assumed that the auditory nerve fibres carried the temporal information present in the acoustic waveform. Much temporal information is indeed carried by the pattern of nerve firings and aspects of this concept still receive support in modern measurements and theories, although each channel must be preceded by its own cochlear filter.
Georg von Békésy (1928 see 1960) was the first to observe the vibration pattern of the cochlear partition. This was mostly done by sprinkling fine silver crystals on Reissner’s membrane and observing the vibration with a stereo microscope using stroboscopic illumination. He made most of these measurements in human cadavers and guinea-pigs, but also in the chicken, mouse, rat, cow and elephant. When plotted as a function of frequency, the results from different specie...
Table of contents
- Cover image
- Title page
- Table of Contents
- ADVANCES IN THE BIOSCIENCES
- Copyright
- Inside Front Cover
- List of Participants
- Preface
- Inner Ear Cell Biology
- Auditory Periphery Functioning
- Frequency Selectivity
- Pitch
- Intensity Coding
- Temporal Analyses
- Spectral Shape Analysis
- Spectro-temporal Processing
- Binaural Interactions And Sound Localization
- Pathologies
- In Honour of Professor Zwicker
- Author Index
- Subject Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Auditory Physiology and Perception by Y. Cazals,K. Horner,L. Demany in PDF and/or ePUB format, as well as other popular books in Medicine & Physiology. We have over 1.5 million books available in our catalogue for you to explore.