
- 312 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Exploring the Thalamus
About this book
The thalamus is a group of cells placed centrally in the brain that serve a critical role in controlling how both sensory and motor signals are passed from one part of the cerebral cortex to another. Essentially, all information reaching the cerebral cortex and thus consciousness is relayed through the thalamus. The role of the thalamus in controlling the flow of information (such as visual, auditory, and motor) to the cortex has only recently begun to be understood. This book provides an in-depth look at the function of the thalamus and its role as relayer of information to the cerebral cortex. The authors explore how the thalamus controls messages that are passed to the cortex and they introduce the novel suggestion that the thalamus serves a critical role in controlling how messages pass from one part of the cortex to another. Exploring the Thalamus is a comprehensive, up-to-date reference for researchers. It discusses problems concerning the function and structure of the thalamus and concludes each chapter with thought-provoking questions regarding future research.
- Focuses on thalamocortical interrelationships
- Discusses important problems concerning the function and structure of the thalamus
- Concludes each chapter with thought-provoking questions requiring future research
Trusted by 375,005 students
Access to over 1 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
Scienze biologicheSubtopic
NeuroscienzaChapter I
Introduction
A Thalamic Functions: What Is the Thalamus and What Is It For?
The thalamus is the major relay to the cerebral cortex. It has been described as the “gateway” to the cortex. Almost everything that we can know about the outside world, or about ourselves, is based on messages that have to pass through the thalamus. It forms a relatively small cell group on each side of the third ventricle and can be seen most readily in a midsagittal section of the brain (Figure 1). It can be divided into several distinct nuclei or nuclear groups, each concerned with transmitting a characteristic type of afferent signal (visual, auditory, somatosensory, cerebellar, etc.) to a cytoarchitectonically distinct and functionally corresponding area or group of areas of the cerebral cortex (Figures 2 and 3) on the same side of the brain. This is the view of the thalamus that was developed during the 70 (plus) years up to circa 1950. It has served us well and is still the view presented in most textbooks. It was based on clinical observations related to postmortem study of the brain and on relatively crude experimental neuroanatomical methods: the Marchi method for staining degenerating myelin in pathways that had been cut or injured, and the Nissl method, which shows nerve cells grouped into functional nuclei or undergoing retrograde degeneration after their axons have been cut. These methods give results in terms of large populations of cells or axons and large areas of thalamus or cortex. Perhaps it was fortunate that modern methods for studying the detailed connectivity patterns of single cells or small groups of cells were not available when the thalamic connections were first being defined. If they had, it is probable that no one would have been able to see the larger thalamic forest for the details of the connectional trees. We shall start with the forest.
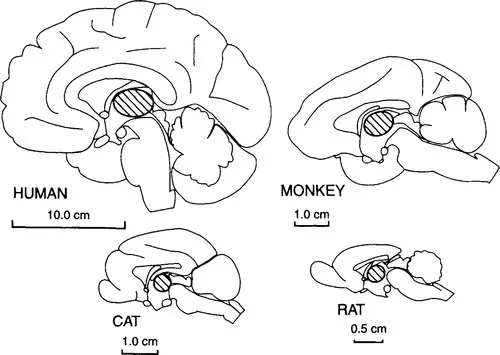
Figure 1 Midsagittal view of the cerebral hemispheres of a human, a monkey, a cat, and a rat (in size order) to show the position and relative size of the thalamus, which is indicated by diagonal hatching.
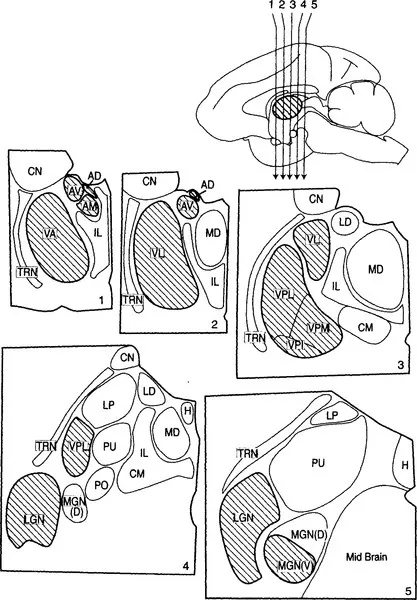
Figure 2 Schematic view of five sections through the thalamus of a monkey. The sections are numbered 1 through 5 and were cut in the coronal planes indicated by the arrows in the upper right midsagittal view of the monkey brain from Figure 1. The major thalamic nuclei in one hemisphere for a generalized primate are shown. The nuclei that are outlined by a heavier line and filled by diagonal hatching are described as first order nuclei (see text), and the major functional connections of these, in terms of their afferent (input) and efferent (output) pathways to cortex, are indicated in Figure 3. AD, anterodorsal nucleus; AM, anteromedial nucleus; AV, anteroventral nucleus; CM, center median nucleus; CN, caudate nucleus (not a part of the thalamus); H, habenular nucleus; IL, intralaminar (and midline) nuclei; LD, lateral dorsal nucleus; LGN, lateral geniculate nucleus; LP, lateral posterior nucleus; MGN, medial geniculate nucleus; PO, posterior nucleus; PU, pulvinar; TRN, thalamic reticular nucleus; VA, ventral anterior nucleus; VL, ventral lateral nucleus; VPI, VPL, VPM, inferior, lateral, and the medial parts of the ventral posterior nucleus or nuclear group.
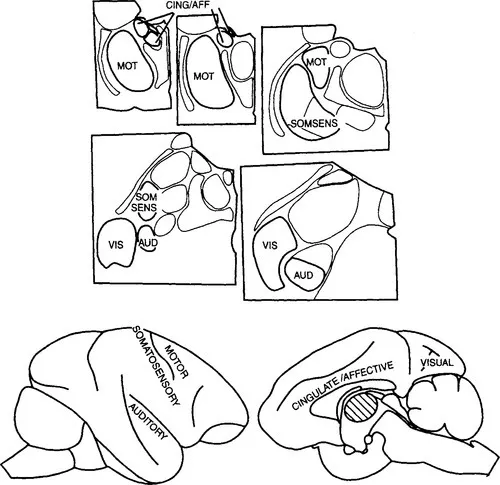
Figure 3 The upper part shows the nuclei illustrated in Figure 2, and the lower part shows a lateral (left) and a medial view of the hemisphere in a monkey to indicate the functional connections of the major first order thalamic nuclei. AUD, auditory; CING/AFF, cingulate/affective; MOT, motor; SOMSENS, somatosensory; VIS, visual.
The schematic view of thalamocortical relationships, summarized in Walker’s great book (1938) or in Le Gros Clark’s earlier review (1932), provided a powerful approach to thalamic function. Even though it was heavily dependent on relatively gross methods, it showed us how to divide up the thalamus and how to relate each of the resulting major thalamic nuclei or nuclear groups to one or another part of the cerebral cortex (see Figures 2 and 3). Above all, it demonstrated the extent to which the functions of any one part of the cerebral cortex must be dependent on its thalamic inputs. It should be noted that the thalamus is connected to the so-called neocortex. Olfactory cortex and hippocampal cortex are not neocortex and do not receive comparable thalamic afferents. Olfactory afferents represent the only pathway of a sensory system that does not have to go through the thalamus before it can reach the cortex. The following account deals with the relationships between thalamus and neocortex only.
Figure 1 shows the thalamus in relation to the rest of the cerebral hemisphere and shows that the thalamus is small relative to the whole cerebral hemisphere in all mammals. There are a great many more cortical cells than there are thalamic cells even though the neocortex depends on the thalamus for its major inputs.1 Each major neocortical area depends on a well-defined thalamic cell group, that is, on a thalamic nucleus or group of nuclei. In the evolutionary history of mammals, an increase in the size of any one part of cortex generally relates to a corresponding increase in the related thalamic nuclei. The functionally best-defined cortical areas (visual, auditory, motor, etc.) depend for their functional specialization on the nature of messages that pass to that cortical area from the thalamus. The visual cortex is visual because it receives visual messages from the retina through its thalamic relay, and the same is true of the other thalamic nuclei outlined in bold and hatched in Figure 2.
Figure 2 shows some of the major thalamic nuclei in a simplified, schematic form for a generalized primate. Details differ for each species, and a number of nuclei are not included in the figure because they play no role or only a minor role in the rest of this book. However, the general relationships shown apply to all mammals. Figure 3 shows how some of these major thalamic nuclei are linked to specific, functionally or structurally defined cortical areas. Further details on individual thalamic nuclei and their connections can be found in Berman and Jones (1982) and Jones (1985). Figures 2 and 3 show that for some, but by no means for all of the thalamic nuclei, we can define the dominant or functionally “driving” afferents. That is, Figure 3 shows that the lateral geniculate nucleus is visual and the medial geniculate nucleus is auditory. The ventral posterior nucleus2 is somatosensory, which is to say that the ascending pathways concerned with tactile stimuli and with stimuli related to the position and movements of body parts (kinesthesis) go to this nucleus as do pathways concerned with pain and temperature. For reasons detailed in Chapter III we treat the afferents from the cerebellum, related to movement control, that go to the ventrolateral and ventral anterior cells groups as drivers and also the mamillothalamic tract that sends information to the anterior thalamic nuclei about ongoing activity in the hippocampal formation and in the midbrain. These represent the major, known ascending driver inputs to the thalamus. We shall treat these ascending afferents as the “drivers” of the thalamic relay cells that they innervate, meaning by this that they are the afferents that determine the receptive field properties of the relay cells. Other afferents, which we shall treat as modulators, can modify the way that the message is transmitted, but they are not responsible for the main qualitative nature of the message that is conveyed to the cortex by the thalamic relay cell. We shall argue that each thalamic nucleus has drivers and modulators and that identifying the drivers for many thalamic nuclei is likely to be a key to understanding their functions, which are currently poorly defined.
The afferents to the other main thalamic nuclei, shown with lighter outlines and no shading in Figure 2 and unlabeled in Figure 3, are less straightforward and will be considered later (Chapters III and VIII). We have presented evidence that these nuclei receive their major driving afferents from the cerebral cortex itself and thus serve as a relay on corticocortical pathways, not as relays of subcortical afferents to cortex (Guillery, 1995; Sherman and Guillery, 1996). That is, they contain “higher order” relays. We define first order relays as those concerned with sending messages to the cortex about what is happening in the subcortical parts of the brain and higher order relays as those that provide a transthalamic relay from one part of cortex to another. It is to be noted that in primates the nuclei that contain the higher order circuits form more than half of the thalamus. How these “indirect” corticocortical relays relate to direct corticocortical connections is a challenging question considered in Chapters VIII and IX.
Although for many of the thalamic nuclei we can show how they serve to connect different cortical areas to sensory surfaces of the body or to other parts of the brain, we cannot readily demonstrate what it is that the thalamus does for the messages that are passed from ascending pathways to the cerebral cortex. Why don’t the ascending pathways go straight to the cortex? This question was always present, but was brought into striking focus in the 1960s when electron microscopists showed the complexities of the synaptic relationships in the thalamus (Szentágothai, 1963; Colonnier and Guillery 1964; Peters and Palay, 1966). Only about 20% of the synapses in the major relay nuclei like the lateral geniculate were then seen to come from the major ascending pathways (Guillery, 1969b), and recent figures show less than half that percentage contact the relay cells that are responsible for passing the information on to the cortex (Van Horn et al., 2000). Complex synaptic formations involving serial synapses and connections from local or distant inhibitory cells characterize all of the thalamic nuclei (e.g., Ralston and Herman, 1969; Jones and Powell, 1969; Morest, 1975), and most thalamic nuclei, in accordance with their shared developmental origin, have more or less the same general organizational plan. Exceptions are considered in later chapters.
The complexity of thalamocortical pathways was further increased by the demonstration of the connections shown in Figure 4. Not only is there a massive input from the deeper layers of the cerebral cortex back to the thalamus, but there is a specialized cell group adjacent to the thalamus, the thalamic reticular nucleus, that receives excitatory branches from the corticothalamic and thalamocortical axons and sends inhibitory axons back to the thalamus (Jones, 1985). The functional role of these reticular connections and of the complex synaptic arrangements found in the thalamus represented (and still represents) a challenging puzzle, a challenge that was greatly increased in recent years by the discovery of diverse transmitters, voltage- and ligand-gated ion channels, and receptors that contribute to the synaptic organization in the thalamic relay (see Sherman and Guillery, 1996; and Chapters V and VI). The functional control of membrane conductances depends on a highly complex interplay of afferent activity and local conditions that will be considered in Chapter IV. These conditions in turn determine the way in which a thalamic cell responds to its inputs and thus determine how messages that come into the thalamus are passed on to cortex. This, the manner in which a thalamic cell passes messages on to cortex, is not constant but depends on the attentive state of the whole animal (waking or sleeping) and probably on the local salience of a particular stimulus or group of stimuli as well: are they new, threatening, interesting, or merely a continuation of prior conditions? (See Chapter VI.)
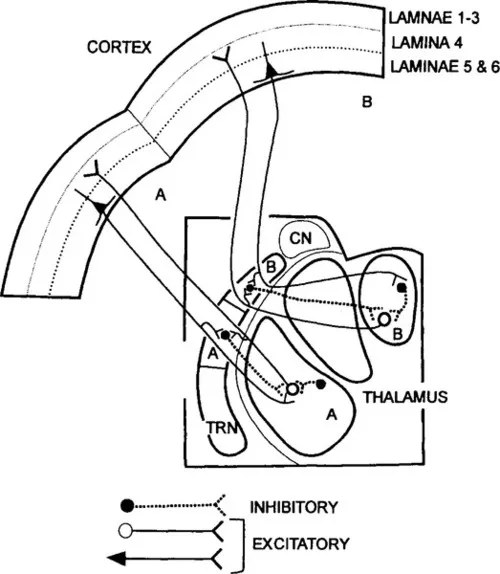
Figure 4 Schematic view of the interconnections between the first order thalamic nuclei, cerebral cortex, and the thalamic reticular nucleus (TRN) as seen on coronal sections of thalamus and cortex. The nuclei labeled A and B are connected with distinct sectors of the reticular nucleus also labeled A and B, and with distinct cortical areas. The connections are shown for components A and B only. The cortical pyramids that give off collateral branches to the reticular nucleus lie in cortical layer 6. CN, caudate nucleus.
When one considers the factors relevant to how the transfer of messages is controlled or gated in the thalamus, it is probable that more than one functionally significant mechanism will be apparent once we have a clear understanding of these aspects of thalamic organization. That is, there are likely to be several more or less distinct functional roles for the synaptic arrangements present in the thalamus. They may be active at different times, or they may have concurrent effects. Two mechanisms that have received significant attention in the recent past have roles in sleep and in the production of epileptic discharges (McCormick and Bal, 1997; Steri...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright page
- Preface
- Abbreviations
- Chapter I: Introduction
- Chapter II: The Nerve Cells of the Thalamus
- Chapter III: The Afferent Axons to the Thalamus
- Chapter IV: Intrinsic Cell Properties
- Chapter V: Synaptic Properties
- Chapter VI: Function of Burst and Tonic Response Modes in the Thalamocortical Relay
- Chapter VII: Maps in the Brain
- Chapter VIII: Two Types of Thalamic Relay
- Chapter IX: Drivers and Modulators
- Chapter X: Overview
- Bibliography
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.4M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Exploring the Thalamus by S. Murray Sherman,Ray W. Guillery in PDF and/or ePUB format, as well as other popular books in Scienze biologiche & Neuroscienza. We have over one million books available in our catalogue for you to explore.