
- 592 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Vascular Transport in Plants
About this book
Vascular Transport in Plants provides an up-to-date synthesis of new research on the biology of long distance transport processes in plants. It is a valuable resource and reference for researchers and graduate level students in physiology, molecular biology, physiology, ecology, ecological physiology, development, and all applied disciplines related to agriculture, horticulture, forestry and biotechnology. The book considers long-distance transport from the perspective of molecular level processes to whole plant function, allowing readers to integrate information relating to vascular transport across multiple scales. The book is unique in presenting xylem and phloem transport processes in plants together in a comparative style that emphasizes the important interactions between these two parallel transport systems.
- Includes 105 exceptional figures
- Discusses xylem and phloem transport in a single volume, highlighting their interactions
- Syntheses of structure, function and biology of vascular transport by leading authorities
- Poses unsolved questions and stimulates future research
- Provides a new conceptual framework for vascular function in plants
Information
Part I
Fundamentals of transport
1
Perspectives on the Biophysics of Xylem Transport
William F. Pickard and Peter J. Melcher
Publisher Summary
This chapter provides a brief discussion on cohesion-tension theory. The chapter also focuses on factors that affect hydraulic resistance in plants. It addresses how sap is extruded into the empty tracheary element even though the free energy gradient from soil to xylem seems often to be in the wrong direction, how the element is hydraulically isolated during refilling to avoid its new contents being sucked away in the transpiration stream, and how a refilled vessel ultimately reestablishes hydraulic contact with neighboring conduits in such fashion as to become once again useful in water transport. For xylem sap to flow through the plant, the negative hydrostatic pressure gradient must be large enough to overcome the “resistive” force of sap viscosity and the force of gravity. Because the rate of xylem sap flow through plants is relatively slow, the laminar flow pattern within an individual vessel consists of layers of sap that flow past each other in a sheetlike fashion, with the result that neither eddies nor vortices occur and turbulence is absent. However, in both laminar and turbulent flow, viscosity leads to internal friction, which produces energy dissipation. The cohesion-tension theory still stands as the hegemonic model for understanding long-distance sap transport in plants, but a controversy related to it propelled the development of new ideas and possible alternative paradigms to replace the postulates of cohesion-tension theory.
“I’m sorry to say that the subject I most disliked was mathematics. I have thought about it. I think the reason was that mathematics leaves no room for argument. If you made a mistake, that was all there was to it.” (X, 1992, p. 35)
The fundamentals of the cohesion-tension theory of sap ascent are now well covered in textbooks (Taiz and Zeiger, 2002; Fisher, 2000) with which most in our potential readership are familiar. Moreover, many deeper questions of xylem biophysics were covered in now hoary reviews (Pickard, 1981; Zimmermann, 1983) and have been updated by Tyree in a substantial monograph (Tyree and Zimmermann, 2002); there is no need to be encyclopedic. Nevertheless, any review of xylem biophysics must include at least some discussion of the cohesion-tension theory. This will be followed by a rather more extensive discussion of how embolisms within the transpiration stream might be formed and, more important, resorbed. Since cohesion-tension theory has only recently survived a significant assault on its hegemony, we present a brief discussion of this controversy, including what has been learned. Finally, we focus on factors that affect hydraulic resistance in plants. In each of these endeavors, some effort will be expended to define each concept with clarity and to frame each explanation rigorously.*
The Biophysics of Sap Ascent in the Xylem
Fundamentals of Cohesion-Tension Theory
In broad outline, the cohesion-tension theory is familiar to virtually every worker in plant biology. We shall therefore treat it as an exercise in dogmatics requiring but little exposition.
Dogma I (Hales, 1727, expt. XXXV):* The rise of the xylem sap during transpiration is due to the transpiration itself, the “capillary… orifices” in the leaves being able “as any sap is evaporated off” to “supply the great quantities of sap drawn off by perspiration” by “their strong attraction (assisted by the genial warmth of the sun).” In modern scientific terminology†: the capillary menisci of the cell walls within the substomatal cavities, being evaporatively depleted by solar heating, contract and draw the sap upwards. Ascribing this core dogma of cohesion-tension theory to Hales is uncommon (but cf. Floto, 1999), it being more usual (e.g., Dixon, 1914; Pickard, 1981) to ascribe it instead to Dixon and Joly (1895) or (e.g., Steudle, 2001) to Böhm (1893).
Dogma II (cf. Dixon, 1914): Water possesses considerable tensile strength, which allows it to be drawn upward by capillary menisci in the leaves. Evidence for this has been reviewed, for example, by Dixon (1914), Pickard (1981), and Steudle (2001).
Dogma III (cf. Haberlandt, 1914): Nevertheless bubbles can form in tracheary elements and interrupt sap flow. This was reported early on (Dixon, 1914; Haberlandt, 1914) and has been much commented upon since (Tyree and Sperry, 1989; McCully, 1999; Canny et al., 2001; Facette et al., 2001).
Dogma IV: Fortunately, plants have developed ways of coping with the challenges posed by emboli. These include (1) anatomical adaptations such as cavitation-resistant conduits and redundancy of hydraulic capacity, (2) new growth, (3) and global pressurization of the xylem as by root pressure. Within the past decade, evidence has appeared for embolus resorption despite the existence of nearby water putatively under tension (e.g., Canny, 1997; McCully et al., 2000; Cochard et al., 2000; Facette et al., 2001; Pickard, 2001; Tyree and Zimmermann, 2002); however, there is as yet no consensus on the precise mechanism(s) by which this might occur (e.g., Hacke and Sperry, 2003; Konrad and Roth-Nebelsick, 2003; Pickard, 2003a; Vesala et al., 2003; Chapter 18, this volume).
The Etiology of an Embolism
Despite the long awareness of tracheary bubbles, there has always been some uncertainty about the mechanism by which they got there, especially after the discovery within the stem of acoustic (Ritman and Milburn, 1988) and electric (Pickard, 2001) transients, which were assumed to arise from putative* cavitation events within the tracheae.
There are four obvious ways by which tension within a liquid-filled pipe could produce an embolism (Pickard, 1981; Zimmermann, 1983; Tyree, 1997).
Homogeneous Nucleation
Consider a spherical bubble of radius r [m] in a liquid of pressure P [Pa] and surface tension γ [N•m−1]; and let the amount of ideal vapor in the bubble be N [mol]. It then follows from the Young-Laplace equation (Pickard, 1981) and the perfect gas law that the pressure differential across the interface is
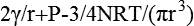
where R [= 8.314 J•mol−1•K−1] is the gas constant and T [K] is the absolute temperature. If the net pressure begins positive, then the bubble will shrink to zero as liquid vapor condenses on its inner surface and other gas is pressurized into solution. If it begins negative and P < 0, then the bubble should expand and fill the available space. What is required is an initial bubble radius such that the net pressure across the interface is negative; and we do not know why that might occur. Detailed theoretical treatments have tradi...
Table of contents
- Cover image
- Title page
- Table of Contents
- Contributors
- Preface
- Acknowledgment
- Part I: Fundamentals of transport
- Part II: Transport attributes of leaves, roots, and fruits
- Part III: Integration of xylem and phloem
- Part IV: Development, structure, and function
- Part V: Limits to long distance transport
- Part VI: Evolution of transport tissues
- Part VII: Synthesis
- Index
- Physiological Ecology
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Vascular Transport in Plants by N. Michelle Holbrook,Maciej A. Zwieniecki in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Physiology. We have over 1.5 million books available in our catalogue for you to explore.