The Antibody Molecule reviews the literature leading to current knowledge of the structure of immunoglobulins. The book begins by outlining some of the basic structural characteristics of immunoglobulins without citing the references on which the information is based. Separate chapters follow covering the chemical nature of the active site of an antibody molecule and mechanisms of interaction with hapten; the general structural features and properties of the various classes of human immunoglobulin; and amino acid sequences of human and mouse L chains and of human and rabbit H chains. Subsequent chapters deal with the evolution of the immunoglobulin classes; special properties of mouse, guinea pig, rabbit, and horse immunoglobulins; idiotypic specificities of immunglobulins; and the genetic control of antibodies. This book is meant for immunologists who have not personally observed the development of this exciting period in the history of immunology. It will also provide useful supplemental reading for the serious student or investigator who wishes to become familiar with the nature of the antibody molecule, its genetic control, and mode of action.
Trusted by 375,005 students
Access to over 1 million titles for a fair monthly price.
General Structural Features of Immunoglobulin Molecules; Myeloma Proteins
Publisher Summary
This chapter discusses the structure of immunoglobulins. Immunoglobulins are present throughout the vertebrate kingdom but have not been identified in invertebrates. A basic feature, maintained during evolution, is the four-chain structure, comprising two light (L) and two heavy (H) chains. Both the H and L chain contribute amino acid residues that form the antigen-binding site. Each H–L pair produces one binding site, and the four-chain structure thus yields two sites. An individual molecule normally contains one species of H chain and one species of L chain, so that the combining sites of a molecule are identical. This property has its origin in the expression of only one gene for an L chain (variable region) and of one gene for the H chain (variable region) in a lymphoid cell that is synthesizing antibody. An immunoglobulin molecule may have more than four chains. Such molecules are polymers of the four-chain unit, which are held together by disulfide bonds.
One purpose of this text is to review the literature leading to our current knowledge of the structure of immunoglobulins. In subsequent chapters, references are provided to much of the pertinent research. In this introductory section we will outline some of the basic structural characteristics of immunoglobulins without citing the references on which the information is based. The discussion will be brief and, at times, sketchy. Its purpose is to facilitate reading of the remainder of the book, and perhaps to permit the reading of other chapters without strict regard to their sequence.
Immunoglobulins are present throughout the vertebrate kingdom but have not been identified in invertebrates. A basic feature, maintained during evolution, is the four-chain structure, comprising two light (L) and two heavy (H) chains. Both the H and L chain contribute amino acid residues which form the antigen-binding site. Each H-L pair produces one binding site, and the four-chain structure thus yields two sites. An individual molecule normally contains one species of H chain and one species of L chain, so that the combining sites of a molecule are identical. This property has its origin in the expression of only one gene for an L chain (variable region) and of one gene for the H chain (variable region) in a lymphoid cell that is synthesizing antibody.
An immunoglobulin molecule may have more than four chains. Such molecules are polymers of the four-chain unit, which are held together by disulfide bonds. For example, molecules of the IgM class may have from 16 to 24 chains (4–6 four-chain units, or 8–12 potential binding sites), depending on the animal species. Serum IgA may also exist as a polymer, and secretory IgA, found in colostrum, tears, saliva, and other secretory fluids, generally occurs as a dimer of the four-chain unit.
Thus the minimum “valence” of an immunoglobulin is 2. The presence of more than one combining site on the antibody molecule is of biological importance because it permits aggregation of macromolecular or particulate antigens. Bivalence is essential for enhancement of phagocytosis by antibody and for fixation of complement in the presence of antigen. Also, bivalent or multivalent attachment of a single antibody molecule to a particle increases the energy of interaction or “avidity”; this is significant, for example, in the neutralization of viruses, particularly when the affinity of a single combining site is low, as is often the case early in the process of immunization.
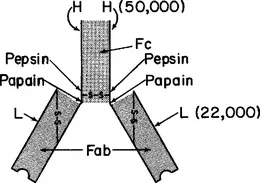
It was demonstrated by R.R. Porter in 1958 that cleavage of rabbit IgG by papain at neutral pH gives rise to large fragments with distinctive properties. The nature of this cleavage, as it applies to rabbit IgG or human IgG1, is illustrated in Figs. 1.1 and 1.2. Papain selectively attacks each H chain at a site just N-terminal to the interheavy chain disulfide bonds. This liberates three large fragments. Two are called Fab fragments (ab for antigen-binding) and the other is fragment Fc (c for crystallizable). Each Fab fragment (mol. wt.
45,000) has one antigen-binding site, i.e., is univalent. Fab fragments therefore cannot precipitate macromolecular antigens or agglutinate particulate antigens, but they can, when present in excess, block the precipitation or agglutination that would otherwise occur upon addition of untreated, bivalent antibody. The fragments do this by occupying antigenic determinants, thus denying access to bivalent antibodies. Each Fab fragment comprises a complete L chain and the N-terminal half of an H chain, designated fragment Fd. The L chain and fragment Fd are held together by noncovalent bonds and by a disulfide bond.
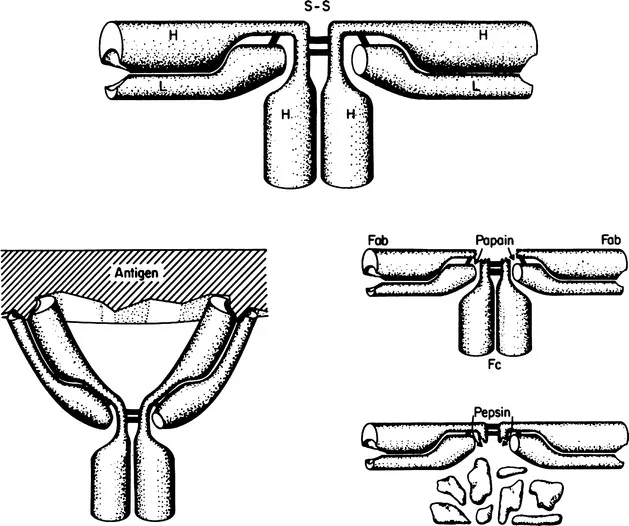
Fig. 1.1 Schematic diagram of the arrangement of polypeptide chains and interchain disulfide bonds in rabbit IgG [after Fleischman et al. (2)]. Reduction of the interchain disulfide bonds does not result in a decrease in molecular weight at neutral pH, owing to noncovalent interactions (shaded regions). A more accurate model would show the two L chains as being in very close proximity in the region of the interheavy chain disulfide bond, since in some immunoglobulins the L chains are disulfide bonded to one another. The molecule is flexible; the Fab fragments can rotate relative to one another around the “hinge region,” which includes the sites of cleavage by papain and pepsin.
Fig. 1.2 Schematic diagram of the arrangement of polypeptide chains in human IgG1.
The third fragment, Fc (mol. wt. 50,000), crystallizes spontaneously in cold neutral buffer from a digest of rabbit IgG, which is heterogeneous. Crystallizability is generally associated with homogeneity and this was the first evidence for the presence of an invariant segment in the heterogeneous IgG molecules. We know now that fragment Fc consists of the C-terminal halves of two H chains, which are held together by noncovalent interactions.
Figures 1.1 and 1.2 also show the site of attack of rabbit or human IgG by pepsin at pH 4 to 4.5. Owing to the fact that its preferential site of cleavage is on the C-terminal, rather than the N-terminal side of the interheavy chain disulfide bond(s), a large bivalent fragment is liberated. Only the interheavy chain disulfide bond(s) join the two univalent fragments after peptic digestion; these bonds are very easily reduced, liberating two univalent fragments, designated Fab′, which are slightly larger than fragment Fab. [The initial product of digestion is F(ab′)2.] The Fab′ segments can be reunited, to form fragment F(ab′)2, by reoxidation of the disulfide bond; this cannot, for obvious reasons, be accomplished with fragment Fab. Pepsin partially degrades fragment Fc, possibly owing to the low pH required for peptic activity.
Because of the susceptibility of a particular region in the middle of the H chain to attack by papain, pepsin, and other enzymes, it is generally thought that this region must be loosely folded. It has been designated the hinge region because the two univalent (Fab) fragments can move in space in relation to one another, and it is believed that the “swivel” is in this region of the chain.
This flexibility permits bivalent attachment of a single antibody to a particulate antigen (horseshoe configuration of the antibody) or, alternatively, the linking of two antigen particles with the antibody stretched out to its maximum length (about 140 Å for IgG). The combining sites are situated at the ends of the bivalent IgG molecule. A single, decavalent IgM molecule can attach itself to a particulate antigen through several combining sites. (Information of this kind is obtained by electron microscopy; the location of the combining site has been confirmed by X-ray crystallography, which also provides information as to the dimensions of the site.)
As indicated in Fig. 1.1, each H chain is joined to an L chain through a disulfide bond. In addition, strong noncovalent bonds serve to link the two chains. The latter can be disrupted by reagents such as 6 M guanidine hydrochloride, 8 M urea, or 1 M propionic acid. If the interchain disulfide bond joining the H to the L chain is first reduced, the chains can be separated on a preparative scale by gel filtration in the solvents mentioned. The chains reassociate as H-L pairs, in quite good yield, if the dissociating solvent is replaced by dialysis against neutral buffer. In the native molecule the H chains are joined to one another through disulfide and noncovalent bonds in the hinge and Fc regions. Under appropriate conditions it is possible to produce half-molecules, each consisting of a complete H chain and a complete L chain.
Besides L and H chains, secretory IgA contains a polypeptide called secretory component, or SC, which has a molecular weight of 65,000 – 70,000 and is bound to the H chains of the molecule. Another component, J chain (J for joining; mol. wt. 15,000), is found in the IgM of many primitive and advanced species, and in polymeric, but not monomeric (four-chain) IgA. J chain is linked to H chains through disulfide bonds. The amino acid sequences and antigenic properties of SC and J chain do not appear to be homologous to those of the immunoglobulin polypeptides; i.e., they do not seem to have a common evolutionary origin.
In humans and in many other species there are two “types” of L chain, κ and λ. The two types have sufficient homology of sequence to indicate a common evolutionary origin: phylogenetic evidence suggests that they both existed prior to the divergence of mammals and birds. It is possible that κ chains arose first in evolution since they appear, on the basis of limited data, to be the only type present in sharks. The two types of chain are approximately equal in length. Each chain has two (sometimes three) intrachain disulfide loops and a half-cystine through which the L chain is disulfide bonded to an H chain; this half-cystine is generally the C-terminal residue in a κ chain and the penultimate residue in a λ chain. The original discovery of κ and λ chains was made with human immunoglobulins, and the designation of a chain from another species as κ or λ is based on similarity of its amino acid sequence to the corresponding human protein. Even among mammalian species the percentages of κ and λ chains vary greatly – from 0 to 100% for each type. Humans have, on the average, 60% κ chains; rabbits, 70–90%; and mice, about 95%; the remainder are λ. When both types are present in a species, κ and λ chains are found in association with H chains of all classes and subclasses.
The size of an L chain is quite invariant (mol. wt. 22,000–23,000) in vertebrates throughout the phylogenetic scale. Also there are strong “homologies” among the amino acid sequences of L chains. (This applies to H chains as well.) In addition, definite homologies are observed when the sequences of H and L chains are compared with one another. This clearly suggests that there was a common ancestral gene for all the immunoglobulins.
The “primeval” polypeptide sequence probably had a molecular weight of 10,000 to 12,000 and comprised about 100–120 amino acid residues. Evidence for this can be seen, for example, in the structure of a typical human L chain, which contains about 214 amino acid residues. For a given type of L chain (κ or λ), the second half of the sequence, comprising about 107 residues at the C-terminal end of the chain, is invariant except for minor, inherited differences, or variants with nearly identical sequences found in all individuals. Wide variations of sequence are seen, however, in the first half (N-terminal 107 residues) of the chain. The sequence variations, in H as well as L chains, account for the differences in specificity among antibodies. It is clear that the L chain can be thought of as comprising two segments, about equal in size. In a...
Table of contents
Cover image
Title page
Table of Contents
IMMUNOLOGY
Copyright
Preface
Chapter 1: General Structural Features of Immunoglobulin Molecules; Myeloma Proteins
Chapter 2: Nature of the Active Site of an Antibody Molecule and the Mechanism of Antibody-Hapten Interactions
Chapter 3: Human Immunoglobulins
Chapter 4: Amino Acid Sequences in Human Immunoglobulins and in Mouse Light Chains
Chapter 5: The Three-Dimensional Structure of Immunoglobulins
Chapter 6: Properties and Interactions of the Light and Heavy Chains of Immunoglobulins
Chapter 7: Evolution of the Immunoglobulins
Chapter 8: Immunoglobulins of the Rabbit, Mouse, Guinea Pig, and Horse
Chapter 9: Allotypes of Rabbit, Human, and Mouse Immunoglobulins
Chapter 10: Homogeneous Antibodies and Myeloma Proteins with Antibody Activity
Chapter 11: Idiotypic Specificities of Immunoglobulins
Chapter 12: Theories of the Genetic Control of Diversity of Antibodies
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.4M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Antibody Molecule by Alfred Nisonoff,John E. Hopper,Susan B. Spring, F. J. Dixon,Henry G. Kunkel in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Zoology. We have over one million books available in our catalogue for you to explore.