
eBook - ePub
¡Hay un dinosaurio en mi sopa!
Una guía rápida sobre evolución biológica
Álvaro Chaos Cador
This is a test
Share book
- 214 pages
- Spanish
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
¡Hay un dinosaurio en mi sopa!
Una guía rápida sobre evolución biológica
Álvaro Chaos Cador
Book details
Book preview
Table of contents
Citations
About This Book
¡Hay un dinosaurio en mi sopa! nos guía por la evolución de los seres vivos con una acertada mezcla de información sobre procesos evolutivos, temas históricos y asuntos culturales. Sus 62 brevísimos capítulos presentan la biología evolutiva para un público amplio, con la precisión del científico profesional a la vez que una mirada propia y fresca, sentido del humor y una prosa que fluye sin obstáculos.
Frequently asked questions
How do I cancel my subscription?
Can/how do I download books?
At the moment all of our mobile-responsive ePub books are available to download via the app. Most of our PDFs are also available to download and we're working on making the final remaining ones downloadable now. Learn more here.
What is the difference between the pricing plans?
Both plans give you full access to the library and all of Perlego’s features. The only differences are the price and subscription period: With the annual plan you’ll save around 30% compared to 12 months on the monthly plan.
What is Perlego?
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 1000+ topics, we’ve got you covered! Learn more here.
Do you support text-to-speech?
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more here.
Is ¡Hay un dinosaurio en mi sopa! an online PDF/ePUB?
Yes, you can access ¡Hay un dinosaurio en mi sopa! by Álvaro Chaos Cador in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Evolution. We have over one million books available in our catalogue for you to explore.
Information
ENREDOS
En el mundo coloquial la palabra red nos remite a dos lugares. Uno se relaciona con ciertos aparejos formados generalmente por hilos o cuerdas cuya finalidad es la de atrapar algo. Las más famosas son las que usan los pescadores para cazar peces. Infortunadamente, en ellas también caen y mueren, en muchas ocasiones, otros animales como delfines, tortugas y tiburones, los cuales, al no ser el objetivo de la pesca, se tiran al mar ya muertos, de la misma forma que se arroja una lata a la basura. Es necesario instrumentar leyes para regular la pesca, pues se cometen grandes ecocidios que a la larga repercutirán en el ecosistema planetario. Las redes son tan eficaces en el agua como en la tierra y el aire. Aves, murciélagos, insectos y bestias caen fácilmente en la trampa. Allá en la Roma imperial era un arma socorrida por gladiadores, aunque, según se cuenta, era difícil ser diestro en su arte. Los humanos tampoco escapan a su eficacia; además de los miles de esclavos que fueron atrapados con ellas, hoy en día, en las ciudades más avanzadas y con mayor poder económico, los hombres siguen cayendo. Allí la estrategia ha cambiado un poco: en lugar de tirar la red sobre el incauto, se mete un par de piernas femeninas en ellas. Con eso basta para atraer a la presa.
La otra connotación familiar de red se refiere a entes interconectados. Es extraño quien no posee una cuenta de Facebook o de Twitter, lugares donde confluyen las personas mediante algún aparato electrónico “inteligente”. Las redes sociales electrónicas se han convertido en un recurso indispensable en la actualidad para mantener contacto rápido e independiente de los oligopolios mediáticos, quienes proveen información sesgada y tendenciosa. Independientemente del fenómeno social, el estudio acerca de cómo se estructuran las redes sociales y de qué manera fluye la información es materia de investigaciones respaldadas por los gobiernos. Un efecto curioso, hasta simpático, es el del mundo pequeño. Cuántas veces habremos oído el dicho “El mundo es un pañuelo” —ojalá que limpio— en referencia a la sorpresa de tener amistades o conocidos mutuos con personas insospechadas. Una red social se urde de manera característica. Tal como sucede con las redes genéticas, hay personas con muchos vínculos, mientras que otras están pobremente enlazadas. En el lenguaje de las redes sociales los nodos se conocen como agentes. Ahora bien, hay dos maneras de estar muy conectado dependiendo del sentido de la flecha. Una persona que conoce a muchas otras, quedará dibujada por un agente del cual emanan múltiples flechas hacia otros agentes, sus conocidos. Sin embargo, que este sujeto conozca a los demás no implica que ellos lo conozcan. El caso invertido es el de un agente al cual llegan muchísimas flechas. Esa persona es muy conocida. Podríamos decir que la primera es una persona muy informada; la segunda, muy popular. Ciertamente es posible que un conocido también sepa quién soy yo. En ese caso la línea que establece la relación se dibuja con flechas en ambos lados. Lo importante es identificar el sentido de las relaciones y tener presente que no son necesariamente simétricas. Un chismoso tiene muchas conexiones hacia fuera —se la pasa parloteando—; un sabio, muchas hacia dentro —todos acudimos por su consejo—. El que me guste cierta chica no se relaciona con que yo le parezca atractivo —situación ampliamente conocida por todos—.
Hace unos años, el actor estadunidense Kevin Bacon se despachó con una frase un tanto chocante. Aseguró que él había trabajado directa o indirectamente con todo el mundo en Hollywood. Por ejemplo, en Apolo 13 compartió créditos con Tom Hanks y Ed Harris. Según Kevin, él está ligado con todos los artistas de la “Meca” del cine. Si usted es un cinéfilo experto, sabrá que Kevin no ha participado en alguna película junto a Penélope Cruz. Pero no crea que ya agarró al actor. Él dijo que su relación era directa o indirecta, con lo cual se refería a una relación no de primer contacto. Si existe algún actor que haya trabajado con Penélope y a la vez con Kevin, la liga existe. Entonces se dice que la distancia es de dos entre el estadunidense y la española. Para investigar este tipo de relaciones se construyó una página en internet bajo el nombre de The Oracle Bacon, en la que se puede conocer el número “Bacon” de cualquier actor. También es posible hallar la distancia entre cualquier par de actores mientras aparezcan en la Base de Datos de Cine en Internet (IMDB, por su sigla en inglés). El número de saltos es pequeño generalmente. Bueno, dirá usted, es que él es muy famoso. No se sienta mal, mi estimado lector. Que Kevin esté ligado a cualquier otro actor por un número reducido de intermediarios no es un fenómeno anormal. La teoría del mundo pequeño propone eso.
El psicólogo estadunidense Stanley Milgram llevó a cabo un experimento interesante en 1967. Dio una carta a varias personas con el remitente al cual deberían de llegar. El destinatario y el emisor fueron escogidos aleatoriamente. Pidió que intentaran hacer llegar la carta a su destino valiéndose de sus conocidos. Según Milgram, todos los humanos en el planeta estamos conectados por no más de seis intermediarios. Gran parte de las cartas llegó a su destino. De allí se reforzó la idea teórica de los seis grados de separación propuesta inicialmente por el húngaro Frigyes Karinthy. En este libro no nos interesa discutir las críticas que se le hicieron al psicólogo, pero sí resaltar la estructura de la red, la cual se asemeja mucho a las genéticas en cuanto a que hay pocos nodos con muchas conexiones y muchos con pocas. Si ansía conocer a alguien, éstas son buenas noticias: necesitará de escasos intermediarios.
Con base en lo anterior, pasemos al modelo de redes genéticas binarias. Dibujemos una red de cuatro nodos: A, B, C y D, unidos de tal manera que A quede ligado con B, B con C, C con D, y D con A. Al final tendremos un cuadrado cuyos vértices son los cuatro genes. La forma de la red se denomina topología. Las ligas son flechas en ambas direcciones. Recordemos que la actividad de un gen depende de uno o más genes con los cuales tiene vínculos; si lo estimula, se dice que es su activador o excitador; si no, es su inhibidor o represor. Y que cuando un gen está activo o encendido se simboliza con un uno; al estar inhibido o apagado, con un cero. Dada la topología de la red, la actividad de A está regulada por B y D. Tanto B como D pueden estar en uno de dos estados: cero (0) o uno (1). Entonces hay cuatro combinaciones de entradas recibidas por A: 00, 01, 10, y 11. Hay que establecer la actividad de A según la actividad de B y D, que es la regla lógica de A. Asignemos la regla Ox, la cual conviene lo siguiente: si B y D están apagados o encendidos ambos, A estará apagado. A se encenderá si B se activa y D no o, al contrario, si D se enciende y B se apaga. Poniéndolo en términos de unos y ceros, la regla es: 00 da 0, 01 da 1, 10 da 1 y 00 da 0. De la misma forma se establece la regla lógica de los demás genes. Es posible que todos tengan la misma o que cada uno posea la suya. Para simplificar el ejemplo, todos tendrán la misma. En teoría, la red puede adoptar 16 combinaciones diferentes —las cuales conforman el congenespacio—. Según la neosíntesis, todas las configuraciones pueden existir en la naturaleza. Escuchemos qué nos dice la autoorganización.
Empecemos con la primera configuración, A = 0, B = 0, C = 0 y D = 0 (0000). Si los cuatro genes están inactivos, ¿puede el sistema permanecer así? Veamos. De acuerdo con la regla Ox, si dos genes están en cero, el gen al cual regulan queda en cero también. El gen A depende de B y D; como ambos están en cero, A queda en cero. Rápidamente podemos ver que lo mismo pasará con los otros tres genes. La configuración 0000 permanecerá así. Se dice que es estable: es un sumidero.
En la segunda configuración (0001), la cosa cambia porque D está encendido. Ahora A recibe cero de B y uno de D, lo cual, según la regla Ox, hace que A se encienda. Para el gen B la situación no cambia, pues tanto A (no se actualiza su estado sino hasta acabar con todos) como C están apagados: sigue apagado. El caso de C es similar al de A. Recibe cero y uno, por lo que se activará. Al finalizar la actualización de los genes, obtenemos 1010. Por lo tanto, la configuración 0001 no es estable.
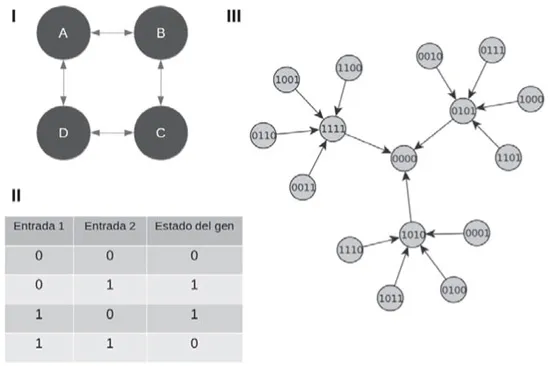
FIGURA 23. I. Topología de la red hipotética de cuatro genes. II. Regla lógica Ox que siguen todos los genes. III. Según la topología y la regla lógica, las 16 configuraciones acaban en 0000. Todas las configuraciones son inestables, excepto 0000, por lo que se le denomina sumidero.
No nos aburriremos evaluando el destino de las restantes configuraciones. En su lugar usaremos un programa computacional para calcularlas.63 Los resultados indican que sólo la configuración 0000 es estable. Las demás no. Se empiece en la que sea, el sistema siempre acabará con los cuatro genes apagados. El modelo predice que de todo el espacio de posibilidades, sólo una de ellas podrá existir, número paupérrimo comparado con el total. Y, además, especifica exactamente cuáles son. Aquel morfoespacio de conchas donde la neosíntesis se aventuraba, llena de inseguridades, para explicar los lugares ocupados y desocupados, cambia drásticamente en el congenespacio y las redes genéticas.
El vastísimo espacio desocupado no puede existir. Esas configuraciones genéticas son inestables. Por más que nos esforcemos en encender los cuatro genes del ejemplo anterior jamás permanecerán así. La misma dinámica del sistema obligará a apagarlos todos. La selección natural podrá elegir entre lo que existe, lo estable; por lo tanto, sólo podrá disponer de individuos donde esa red tenga el cuarteto de genes inactivos.
Si bien los modelos nos transportan al mundo de la imaginación y de la manipulación absoluta, donde podemos jugar a ser constructores y diseñadores de mundos, siempre rogamos por poder concretar esas elucubraciones en situaciones reales para probarlos. En tanto no pase eso, quedan como ciencia ficción. Lo bonito del modelo de redes genéticas binarias es que ha podido ser validado con datos experimentales, es decir, reales. Sin duda, el estudio más bonito de este tipo involucra la floración de un vegetal.
La arabidopsis es una planta diminuta. Posee flores chiquitas, de acuerdo con su tamaño. Son blancas. La importancia de ella reposa en que su genoma también es chico, por lo cual se utiliza comúnmente en la biología para hacer experimentos. Es obvia la ventaja de trabajar con sistemas reducidos. Establece el punto de partida para entender procesos biológicos en los vegetales. Por eso se le llama planta modelo. Desde siempre, la flor ha sido un objeto emotivo para el enamorado y enigmático para el científico. Así como el surgimiento de la reproducción sexual no está muy claro científicamente, el origen de la flor, menos. El mismo Darwin apelaba como “el misterio abominable” a la formación de la estructura vegetal. Actualmente se han descrito varios modelos basados en datos experimentales sobre cómo la interacción de ciertos genes provoca su génesis. Está bien entendido, bajo el modelo ABC, qué tipo de combinaciones genéticas producen las diferentes partes que componen una flor. Sin embargo, dichos modelos, más que predecir son explicadores. El grado de madurez de una ciencia cuyo poder sea la predicción es superior al de otra donde nada más se narran los sucesos. Generalmente, durante el proceso de desarrollo científico, el primer paso es describir, para luego aventurar hipótesis y, finalmente, cuando se comprueban, predecir el fenómeno natural.
El modelo ABC es narrativo. Con él se describen cuáles son los genes que deben de activarse e inactivarse para producir la cuarteta de verticilos florales: sépalos, pétalos, estambres y carpelos. Primero identifica tres tipos de genes: los A, los B y los C. Explica que la formación de los sépalos depende solamente de la actividad de los genes A. Los pétalos necesitan, para producirse, la acción de los genes A y los B exclusivamente. Para que se formen los carpelos deben de encenderse nada más los B y los C. Por último, cuando los C son los únicos prendidos, se forman los carpelos. Se ha detectado que cada una de estas estructuras florales surge de una parte cada vez más interna del tallo. Los sépalos son los más externos, luego vienen los pétalos, posteriormente los estambres y, al final, los carpelos (sépalo, pétalo, estambre y carpelo). Gráficamente se representa con cuatro circunferencias concéntricas, cada una más pequeña que la anterior, detallando un corte transversal al tallo, como puedes observar en la figura 24, donde la circunferencia más externa delimita la influencia de los genes A; la siguiente, la de los A y B; la tercera, la de los B y C, y la más interna, la de los C. El modelo también representa los mutantes. Si cae una mutación que descompone los genes A, la zona donde actuaban queda vacía, entonces los B la ocupan. El resultado es un mutante A: una flor sin sépalos y sin pétalos, pues recordemos que su formación requiere genes A. Ahora los C ocupan la zona de los A, causando el desarrollo de carpelos donde antes había sépalos y estambres en el lugar de los pétalos. En las zonas circulares más internas, la tercera y la cuarta, no hay cambios. El arreglo floral del mutante A es carpelo, estambre, estambre y carpelo. Al mutar los genes B se forma un mutante B cuya flor tendrá sépalos en los dos primeros verticilos, y carpelos en los dos últimos. Quedando el arreglo floral como sépalo, sépalo, carpelo y carpelo, puesto que los genes A y C son los que están activos. Para finalizar, los mutantes C presentan sépalos y pétalos en las posiciones normales, pero donde deberían de aparecer los estambres y los carpelos salen pétalos y sépalos nuevamente. Su orden es sépalos, pétalos, pétalos y sépalos.
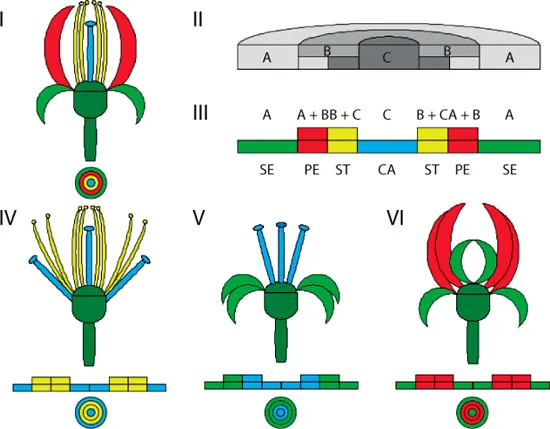
FIGURA 24. En todas las figuras el verde claro simboliza los sépalos; el rojo, los pétalos; el amarillo, los estambres, y, finalmente, el azul, los carpelos. I. Arriba: Esquema de una flor. Abajo: Sección transversal del tallo detallando las zonas por donde surgen los diferentes verticilos de la flor en condiciones normales: sépalos, pétalos, estambres y carpelos, en orden de afuera hacia dentro. II. Corte transversal del tallo mostrando las áreas de influencia de los genes A, B y C. III. Modelo esquemático del corte anterior. IV. Mutante A, donde se pierden los sépalos y los pétalos. Se presenta cómo es su flor y la influencia de los genes B y C en las áreas del tallo. V. Mutante B (ib). VI. Mutante C (ib).
Un equipo de investigadores mexicanos decidió utilizar el modelo de redes genéticas binarias para explicar la formación de la flor. Partiero...