
eBook - ePub
Máquinas vivientes
¿Cómo se mueven las células?
Isaura Meza, Eugenio Frixione
This is a test
Share book
- 141 pages
- Spanish
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Máquinas vivientes
¿Cómo se mueven las células?
Isaura Meza, Eugenio Frixione
Book details
Book preview
Table of contents
Citations
About This Book
El movimiento es uno de los fenómenos que más han llamado la atención de quienes se han dedicado a observar y estudiar la naturaleza. ¿Cómo se organiza una célula o un conjunto de células para producir movimiento? En este libro se describe cómo se fabrican las diferentes partes de la maquinaria celular a partir de instrucciones contenidas en el material genético.
Frequently asked questions
How do I cancel my subscription?
Can/how do I download books?
At the moment all of our mobile-responsive ePub books are available to download via the app. Most of our PDFs are also available to download and we're working on making the final remaining ones downloadable now. Learn more here.
What is the difference between the pricing plans?
Both plans give you full access to the library and all of Perlego’s features. The only differences are the price and subscription period: With the annual plan you’ll save around 30% compared to 12 months on the monthly plan.
What is Perlego?
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 1000+ topics, we’ve got you covered! Learn more here.
Do you support text-to-speech?
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more here.
Is Máquinas vivientes an online PDF/ePUB?
Yes, you can access Máquinas vivientes by Isaura Meza, Eugenio Frixione in PDF and/or ePUB format, as well as other popular books in Ciencias biológicas & Biología celular. We have over one million books available in our catalogue for you to explore.
Information
Topic
Ciencias biológicasSubtopic
Biología celularV. Motores y máquinas simples: mecanoenzimas y polímeros de soporte
Para proceder en el orden cronológico de sus respectivos descubrimientos, entre los motores celulares debe mencionarse, en primer lugar, a la miosina, la cual se describió e ilustró en capítulos anteriores.
MIOSINA
La miosina, con sus dos cabezas en uno de sus extremos, tiende espontáneamente a formar fascículos con un arreglo peculiar. Cada molécula se une a otra con orientación opuesta, empalmando las colas, y cada una de estas parejas se asocia lateralmente con otras parejas hasta constituir fibras en donde las cabezas quedan asomando escalonadamente en la periferia del fascículo. Constituida de esta manera, la apariencia de la fibra es similar a la que presentaría un ramo doble de rosas en botón, en el que las varas sin hojas estarían empalmadas en sentido contrario, y los botones quedarían expuestos alternada y simétricamente a lo largo de los extremos. Las cabezas de miosina son enzimas capaces de hidrolizar ATP y aprovechar la energía liberada (Figura V.1). La miosina utiliza esta energía para modificar de manera transitoria su propia conformación, y puesto que este cambio produce trabajo mecánico, como veremos a continuación, se trata de una mecanoenzima.
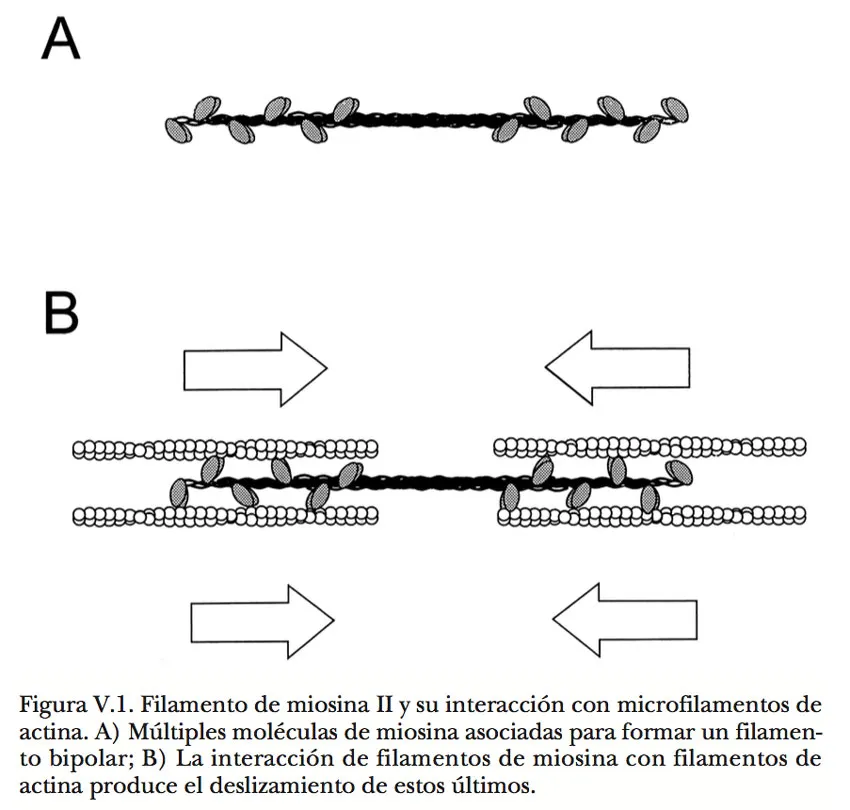
Cuando una de estas fibras se encuentra con un microfilamento alineado en paralelo, las cabezas de miosina propenden a acoplarse con las moléculas de actina, favoreciendo una firme asociación entre ambas estructuras. Es entonces cuando, en presencia de ATP, se expresan las propiedades motoras de la miosina y sus cabezas literalmente caminan a lo largo del filamento. Al igual que el movimiento de un pie, este notable fenómeno ocurre en varias etapas: 1) separación del punto de contacto inicial; 2) avance hacia una nueva posición; 3) afianzamiento en el nuevo punto de contacto, y 4) efecto de arrastre sobre el resto de la estructura en esa dirección. Las moléculas de ATP participan en las dos primeras etapas a través de dos acciones diferentes. Por una parte, tienen la virtud de desprender las cabezas de miosina del filamento de actina; por la otra, ceden su energía para permitir que las cabezas ya liberadas se enderecen de su condición retraída normal y puedan alcanzar el siguiente punto de anclaje. Así, cada cabeza de miosina se suelta del filamento al pescar una molécula de ATP, de éste toma energía para enderezarse, y se ancla nuevamente en un punto vecino del filamento. Al concluir este último paso, la cabeza de miosina se vuelve a retraer y, puesto que se halla unida al filamento de actina, la retracción se refleja en el arrastre de su cola (Figura V.2). El ciclo se repite una y otra vez, siempre que haya ATP disponible. Una cabeza de miosina consume la energía que aportan entre 5 y 10 moléculas de ATP cada segundo para desarrollar este trabajo. Dado que otras cabezas de miosina en esa misma fibra efectúan un proceso equivalente, la fibra se desplaza longitudinalmente sobre el filamento.
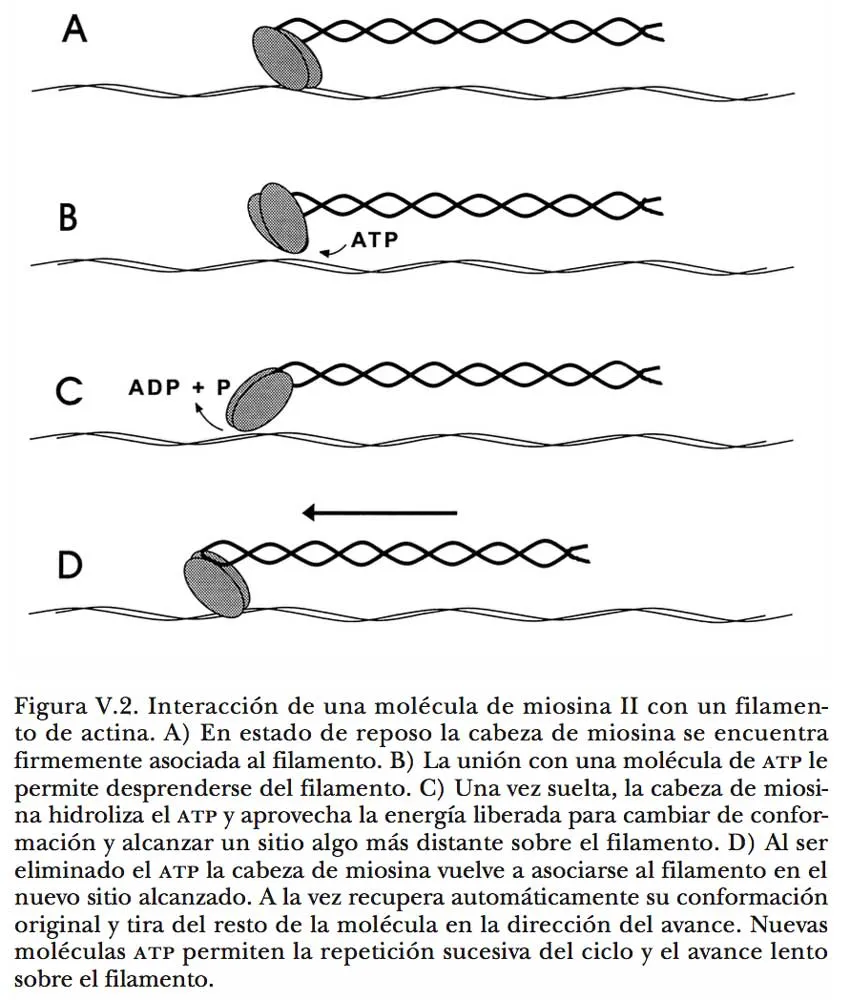
Esta descripción del funcionamiento de un motor molecular no es producto de la imaginación. Aunque no es posible ver directamente la interacción dinámica entre las moléculas de actina y miosina, existen numerosos indicios de que así es como opera, y hace algunos años se obtuvo una prueba contundente de ello. Los investigadores purificaron moléculas de miosina a partir de músculo de conejo, y las fijaron por métodos químicos sobre esférulas de un material sintético visible al microscopio. Cuando estas esférulas se pusieron en contacto con los filamentos de actina que existen en ciertas algas verdes, de inmediato se observó que migraban a lo largo de los filamentos, y el movimiento sólo ocurrió si existía ATP en el medio. Además de corroborar la teoría formulada previamente acerca del mecanismo de deslizamiento de fibras de miosina sobre filamentos de actina, este ingenioso experimento puso en evidencia que dicho mecanismo es común en células tan distintas y distantes entre sí, desde el punto de vista evolutivo, como son las del músculo de un animal superior y las de una planta primitiva. Se trata, por consiguiente, de un recurso tan eficaz que la evolución lo ha conservado y distribuido entre una gran variedad de organismos para desempeñar múltiples funciones.
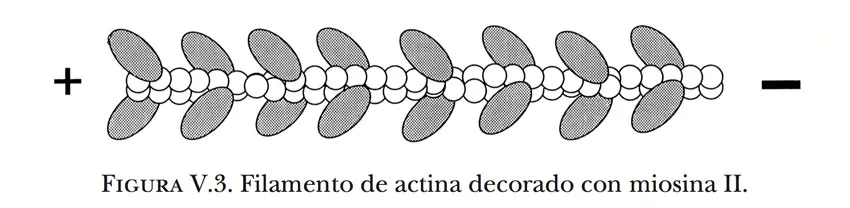
Un filamento de actina se convierte así en una pista para que la miosina camine sobre él, y esto impone una orientación definida al movimiento en el espacio tridimensional. Además, tal como sucede con la mayoría de las pistas, aparte de que el desplazamiento debe efectuarse a lo largo del filamento, la dirección está también determinada: las cabezas de miosina pueden caminar únicamente en un sentido. Esto es consecuencia de la polaridad estructural de los filamentos de actina, que acepta la asociación entre ambas clases de moléculas exclusivamente en una posición específica, de manera análoga o como los asientos de un avión determinan una posición regular y orientada de los pasajeros. Esta propiedad puede demostrarse luego de exponer filamentos desnudos ante una multitud de cabezas de miosina dispersas en una solución carente de ATP. En tales condiciones, por lo explicado antes, la miosina se adhiere con firmeza a los filamentos, y el microscopio electrónico revela que la adhesión ocurre de acuerdo con un arreglo preciso. Las cabezas aparecen recubriendo por completo cada filamento, ordenadas una junto a otra como pequeñas salientes inclinadas que dan al conjunto el aspecto de una apretada serie de puntas de flecha, similar al de una espiga de trigo (Figura V.3). Se dice entonces que el filamento fue decorado con las cabezas de miosina, y ello constituye una prueba contundente de que se trata de un filamento de actina. Cuando las cabezas forman parte de una fibra de miosina en la condición natural, el desplazamiento de la fibra es en dirección contraria a aquella en la que señalan las puntas de una flecha. En términos de la polaridad del filamento de la actina, esta dirección es la que ve hacia el extremo (+), o aquél en donde ocurre preferentemente la adición de moléculas de actina durante la polimerización.
La explicación de los mecanismos que median la interacción motora de la miosina con los filamentos de actina ha sido un camino arduo en el que han participado muchos brillantes investigadores, y dista aún de completarse en todos sus detalles. Sin embargo, por compleja que pueda parecer la descripción dada en los párrafos anteriores, el principio de operación es, en última instancia, el de una de las máquinas más simples inventadas por el hombre; corresponde a lo que en mecánica se llama un trinquete, es decir, un dispositivo en el que una uñeta avanza a lo largo de un poste, clavándose sucesivamente en ranuras hendidas de manera periódica sobre la superficie del mismo. En cada paso, la uñeta se separa de la ranura que ocupa y alcanza la ranura siguiente, donde se engancha para iniciar un nuevo ciclo. La forma de la uñeta y las ranuras es complementaria y oblicua con respecto al eje del poste, de tal modo que el avance sólo puede efectuarse en una dirección; el mecanismo se atranca en la dirección contraria. De manera análoga, la miosina avanza unidireccionalmente como una uñeta sobre el filamento de la actina. La única diferencia fundamental es que la uñeta del trinquete mecánico responde pasivamente a una fuerza externa —aplicada por la mano de un operador, por ejemplo—, mientras que la miosina consume energía directamente para realizar ella misma todo el trabajo. La energía obtenida del ATP es utilizada por la miosina de manera específica para enderezar la cabeza y ponerla en posición de alcanzar un nuevo sitio de anclaje. Es por esta razón que la miosina, a diferencia del trinquete, es un motor.
Dado que las cabezas de miosina deben tener una orientación particular para asociarse funcionalmente con los filamentos de actina, una fibra de miosina, organizada simétricamente con cabezas caminantes apuntando en direcciones opuestas, sólo puede interaccionar con filamentos orientados de manera adecuada en cada uno de sus dos extremos. Sin embargo, el extremo opuesto puede interaccionar con filamentos de actina colocados en posición antiparalela, es decir, cuya polaridad esté invertida con respecto a los primeros. La actividad motora de la miosina en ambos extremos de la fibra resultará entonces en una aproximación de los dos conjuntos de filamentos de actina, si éstos se encuentran libres. Pero si, como es habitual, los filamentos se encuentran unidos en sus cabos distantes con membranas u otros componentes celulares, tales componentes experimentarán una fuerza de tracción hacia la zona en que se halla la miosina. Puesto que las fibras de miosina tienen cabezas caminantes proyectadas en diversos ángulos en torno a su eje, cada fibra puede interaccionar a la vez con varios filamentos de actina en cada extremo. Se multiplica así el trabajo realizado por la fibra y aumenta la fuerza que es capaz de generar. Éste es, esencialmente, el fenómeno que ocurre durante la contracción muscular aunque, como veremos en capítulos posteriores, las células aprovechan este mecanismo para diversos fines. La combinación de este motor molecular —la miosina— con filamentos que transmiten la fuerza constituye una de las máquinas biológicas más ubicuas y, por consiguiente, más estudiadas y mejor comprendidas. Veamos ahora otros ejemplos.
DINAÍNA, CINESINA Y DINAMINA
El funcionamiento del complejo actina-miosina hace patente que las estructuras del citoesqueleto resultan ideales como puntos de apoyo, guías y líneas de transmisión de las fuerzas desarrolladas por los motores moleculares en las células. No sorprende, por tanto, que los microtúbulos, algo más rígidos y resistentes que los filamentos de actina, constituyan el soporte para la operación de otros motores moleculares. Y en retrospectiva, tampoco causa mucho asombro que tales motores estén diseñados conforme al mismo principio que la miosina. De hecho, al menos en el primer caso que abordaremos a continuación, existe una gran similitud (Tabla 1).

La dinaína —la proteína de la fuerza— es otra mecanoenzima que utiliza energía derivada de la hidrólisis del ATP para cambiar transitoriamente su conformación. Se trata también de una molécula de grandes dimensiones integrada por varias subunidades, las mayores de las cuales son dos cabezas globulares con un peso de 410 000 daltones cada una, en las que reside la actividad de ATPasa. Esta actividad aumenta sustancialmente en presencia de microtúbulos. La dinaína fue inicialmente identificada como el motor que genera el movimiento de apéndices vibrátiles llamados cilios y flagelos que ciertas células poseen. En fechas más recientes ha podido comprobarse que hay otras formas de dinaína distribuidas en el citoplasma de una vasta variedad de organismos
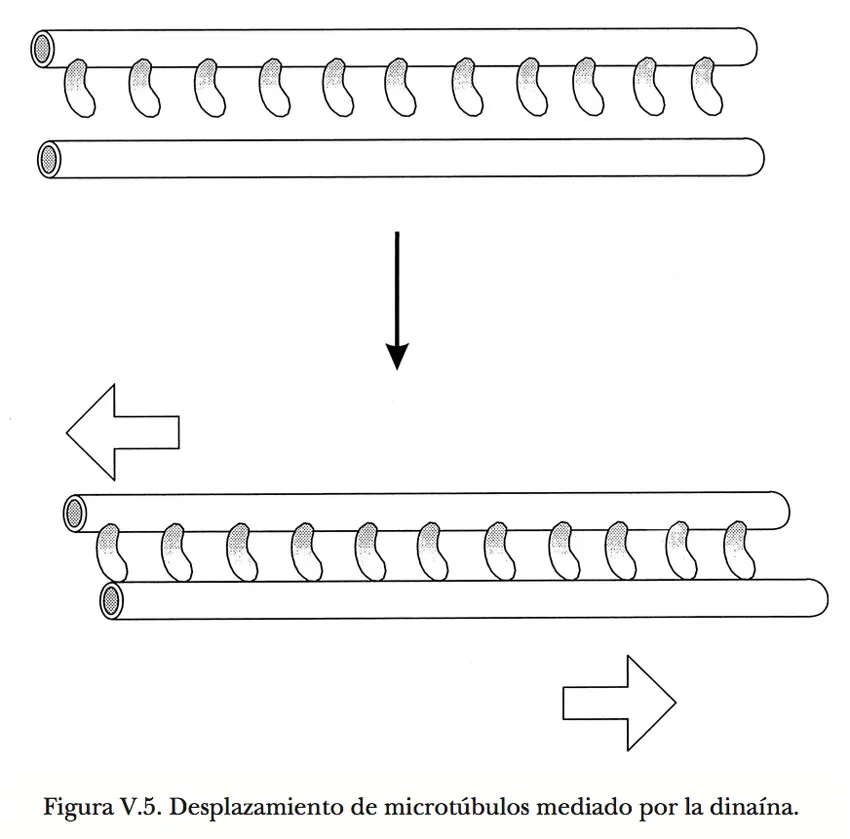
La característica distintiva de las dinaínas es su capacidad para avanzar sobre los microtúbulos, de manera análoga a como lo hace la miosina sobre los filamentos de actina (Figura V.4). Cabe aclarar que esta interacción es por el exterior y no por la estrecha luz de los microtúbulos, como justificadamente alguien pudiera pensar. Todo indica que las cabezas de la dinaína se afianzan a un sitio en la pared de los microtúbulos, en seguida cambian bruscamente de forma, y luego se desprenden para enderezarse y afianzarse en un nuevo sitio, repitiendo el ciclo sin cesar mientras dispongan de ATP. Puesto que normalmente las moléculas de dinaína están unidas a estructuras celulares por el extremo opuesto al de las cabezas motrices, dichas estructuras responden al esfuerzo de arrastre que se ejerce sobre ellas. Cuando la dinaína se halla asociada con una vesícula o algún otro tipo de partícula en libertad para desplazarse, ésta migra a lo largo del microtúbulo. Pero si la dinaína se encuentra anclada en otro microtúbulo, se produce un deslizamiento de un microtúbulo sobre el otro y cada uno de ellos actúa como elemento de transmisión de la fuerza generada hacia las partes de la célula con las que esté conectado mecánicamente. El fenómeno puede reproducirse experimentalmente colocando microtúbulos purificados sobre una laminilla de vidrio recubierta con moléculas de dinaína; se observa entonces que los microtúbulos se deslizan paralelamente a la superficie si el medio está provisto de ATP.
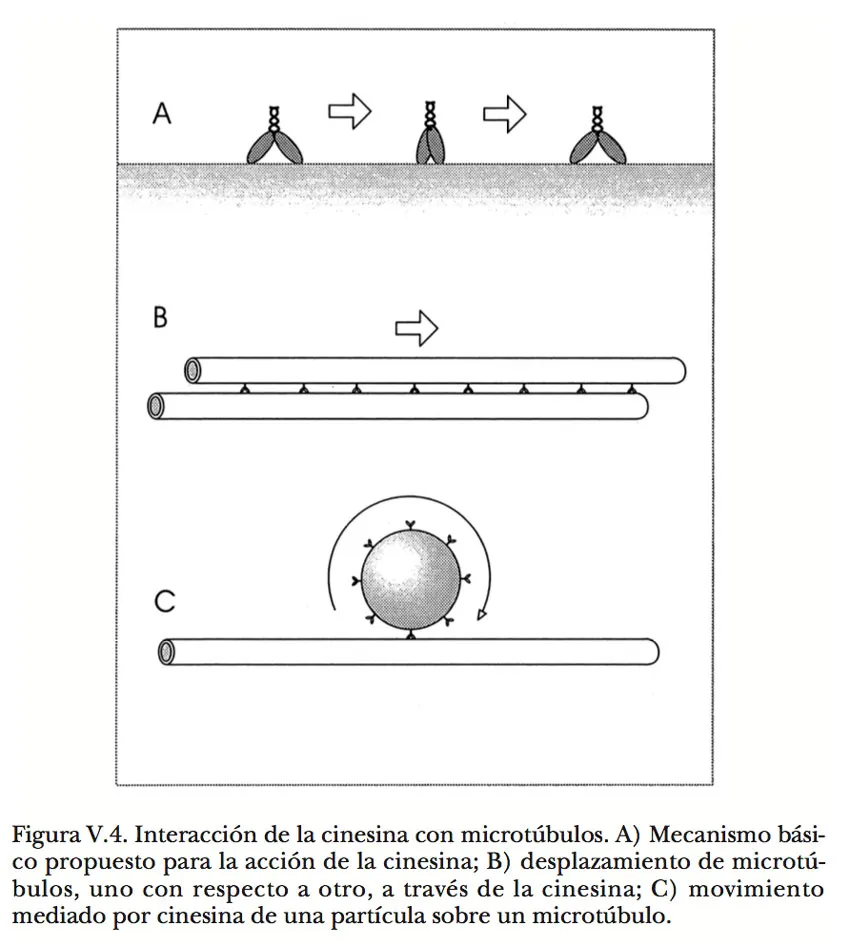
Al igual que en la máquina constituida por actina y miosina, la polaridad de los microtúbulos determina la dirección del avance de la dinaína sobre ellos. Una diferencia importante, sin embargo, es que en este caso el sentido del movimiento apunta hacia el extremo (-) del microtúbulo, es decir, el menos propicio para la adición de moléculas de tubulina durante su polimerización. Por fortuna, en los últimos diez años han aparecido otros dos tipos de mecanoenzimas —las cinesinas o proteínas del movimiento (120 000 daltones) y la dinamina o proteína de la potencia (100 000 daltones)— con propiedades y funciones semejantes a las de las dinaínas, pero que se trasladan en sentido contrario al de éstas sobre los microtúbulos (Figura V.5). En consecuencia, un mismo microtúbulo puede servir como soporte y guía para movimientos de otros componentes celulares en ambas direcciones.
Hemos completado así un inventario mínimo de partes e ingredientes indispensables para la integ...