Discussing the systemic immune response in the contexts of health, disease, and therapy, this unique resource-the only broadly based book of its kind available on the subject-offers comprehensive examinations of the pathways and agents that affect the human immune response and provides state-of-the-art presentations on practical methods of immune modulation.
Focuses on the immune response and modulation in infectious diseases, such as HIV, hepatitis, and parasitic infections and highlights immune modulating agents in gastrointestinal diseases, sepsis, cancer, and autoimmunity!
Written by over 50 international authorities representing distinguished institutions in nine countries, Immune Modulating Agents

- 576 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Immune Modulating Agents
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
MedicineSubtopic
Immunology1
The Immune System and Immune Modulation
Centre National de la Recherche Scientiftque, Villejuif France
I. INTRODUCTION
It is generally agreed that the concept of immune modulation emerged in 1796, when Edward Jenner succeeded in protecting people against smallpox by infecting them with cowpox virus. Since then, many attempts have been undertaken to help the immune system to face external attacks such as bacteria or viruses and internal attacks such as cancers or autoimmune disorders. It is particularly during the past two decades that information on how to manipulate immune responses has been accumulating. The first breakthrough came from the elucidation of the primary structure of the antigen receptor (antibodies and T cell receptors) and the way their diversity is generated. The description of how immune reactions can be elicited or aborted, the insight mechanisms allowing self-tolerization (in particular deletion and anergy), and the discovery of classes of response (for instance, the T helper I [Th1] and Th2 responses) have provided the conceptual bases of interventions on the immune system. Other innovations in immune modulation became possible when the concept of cellular “cross-talk” arose. The discovery of cytokines, capable of influencing the fate of immune cells over some distance, was a major breakthrough to the understanding of how immune responses could be coordinated in the body. Moreover, it is now well established that immune cells, in order to function properly, must cooperate via direct cell-to-cell contacts. For the purpose of immune modulation, it is also important that some open-minded scientists conceived that the immune system does not function in a strictly autarchic manner but rather as an open integrated system. Immune cells, for instance, continually receive inputs from the neuroendocrine system and in turn produce factors that can modulate neural and endocrine responses.
In this chapter, we will outline some general rules governing the immune response. We then will provide an overview on communication within the immune system and between the immune system and other communication systems. On the basis of these premises, we will discuss the possible levels of immunomodulatory interventions. As a cautionary note, this survey does not pretend to be exhaustive, but rather to appeal to the reader’s curiosity, which will, it is hoped, be satisfied by the subsequent chapters of this handbook.
II. OVERALL PRINCIPLES OF IMMUNE REGULATION
A. Specific Versus Nonspecific Signals
Lymphocytes expressing clonally distributed antigen receptors at the cell surface constitute the principal elements of the immune system. Cognate activation of these clonally distributed polymorphic receptors with their physiological ligands (peptide/major histocompatibility complex [MHC] complexes in the case of the T cell receptor, antigen in the case of suface immunoglobulin) is specific in the sense that it only concerns a minor fraction of the overall immune repertoire. In addition, lymphocytes receive signals via monomorphic receptors (coreceptors, cytokine receptors). Such noncognate signals are nonspecific inasmuch as they can be received by the majority or a large fraction of lymphocytes. In this sense, immunomodulatory interventions on monomorphic receptors are always nonspecific (Table 1). Molecules that are designed to interact with antigen receptors also can vary in their specificity. Thus, for instance, antibodies that recognize common features of the T lymphocytes bearing αβ-type antigen receptor/cluster designation (TCRα/β/CD3) complex will have nonspecific effects, whereas antigenic peptides will have strictly specific effects. On an intermediate stage, reagents that recognize a substantial portion of the immune repertoire (e.g., superantigens or antibodies recognizing products from V genes of the TCR or immunoglobulin [Ig] loci) can be viewed as semispecific modulators (Table 1).
| Degree of specificity | Definition | Examplesa |
|---|---|---|
| | ||
| Specific intervention | Modulation of monoclonal or oligoclonal response to a determined antigen | Immunization with pep-tides or proteins; application of modified antigens |
| Semispecific intervention | Intervention on a fraction of the immune repertoire determined by the expression of determined V genes | Superantigens. TCR-V/β or TCR-Vα-specific antibodies, and immunotox-ins. T cell vaccination |
| Nonspecific intervention | Application of substances that alter immune responses independently from the antigen or antigen receptor(s) involved | Application of cytokines, hormones; ablation of T cell subsets |
a TCR; T cell receptor.
B. Three Existential Choices
Both T and B lymphocytes are constantly challenged with three principal choices: (1) survival versus death, (2) response versus anergy, and (3) different classes of response (Figure 1).
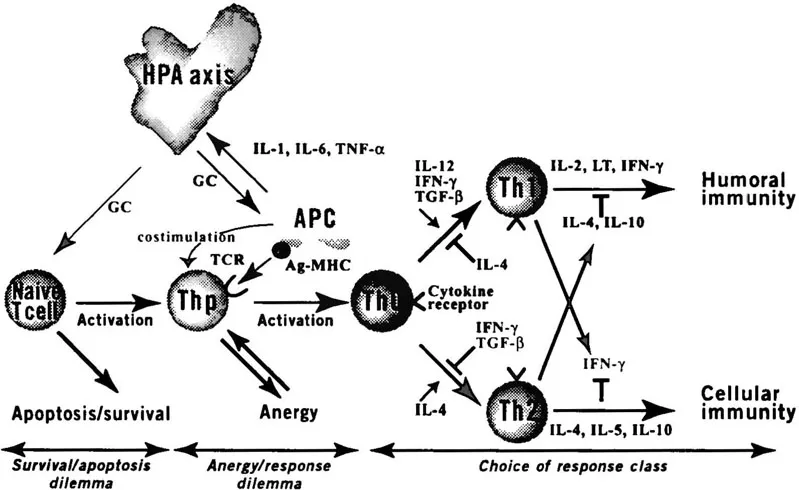
Figure 1 Schematic representation of the immune system showing the different levels of possible immune modulation. Naive peripheral T cells can be activated either to differentiate into a T helper precursor or to undergo programmed cell death (apoptosis). Moreover, activation by antigen presenting cells does not imply productive responses by the T cell. Depending on the simultanous provision of costimuli, the T cell can become anergic or respond in a productive fashion. At a given point, CD4+ T cells “decide” between at least two options, namely, to acquire a Th1 or a Th2 phenotype. This decision process is determined by the type of costimuli provided by APCs as well as the dominant cytokine profile in the environment of the T cell. Th1 and Th2 cells are mutually antagonistic. Thus, for example, Th1 cytokines (especially IFN-γ) favor the differentiation from the Th0 to the Th1 phenotype but inhibit that of Th0 to Th2 cells. In contrast, the Th2 product IL-4 has an opposite effect. It would be an oversimplification to assume that T cells are sequentially exposed to the survival/apoptosis, anergy/response, and class of response dilemmas. Instead, it is conceivable that T cells can be induced to undergo apoptosis or to enter a state of anergy at any differentiation stage. In the upper part of the figure, one particular type of neuroendocrine immune control is depicted. APC can produce inflammatory cytokines (IL-1, IL-6, TNF) that will cause an endocrine arousal reaction and ultimately an increase in adrenal glucocorticoid release. Glucocorticoids then function as endogenous immunosuppressors and exert multiple inhibitory effects, at the levels of both the APC and the T cell. APC, antigen presenting cell; GC, glucocorticoids; HPA axis, hypothalamic-pituitary-adrenal axis; IFN-γ, interferon-γ; IL, interleukin; Thp, CD4+ T helper precursor lymphocyte; TGF-β, transforming growth factor β; TNF, tumor necrosis factor.
At a first level, T and B lymphocytes are notoriously prone to deletion, both during their differentiation in the thymus or bone marrow, respectively, and later during their life in peripheral lymphoid organs. Deletion of lymphocytes results from programmed cell death (apoptosis) and can be achieved by numerous adverse conditions: ligation of “death receptors” (Fas/apoptosis [APO]-l/CD95, CD30, tumor necrosis factor receptor (TNF-R), cytotoxic T lymphocyte antigen (CTLA)-4 on T cells; Fas/APO-l/CD95 on B cells) via cell-to cell contact, presence of soluble death-promoting factors (glucocorticoids tumor necrosis factor [TNF], transforming growth factor-β [TGF-β]), unfavorable combinations of signals, or simply absence of obligatory trophic factors [1,2]. It appears nowadays that T and B cells can be driven into apoptosis at any differentiation stage. Thus, the dilemma of death versus survival constitutes one primary level of endogenous immune regulation.
At a second level, surviving lymphocytes that are activated by nominal antigen can respond in two opposite fashions. On the one hand, lymphocytes can receive the entire set of signals required to mount a productive immune response and thus exert cytotoxic functions to destroy the antigen-bearing target or secrete soluble effector molecules (cytokines in the case of T cells, antibodies in the case of B cells). On the other hand, failure to provide certain signals, the so-called costimuli, can abort the immune response and cause anergy, namely, a reversible partial loss of cellular functions [3,4]. Anergic cells thus, instead of becoming deleted, will fail to exert effector functions, although they may conserve other functions such as self-tolerization.
At a third level, surviving nonanergic lymphocytes can mount different classes of responses. Thus, it is established that undifferentiated CD4+ T cells can become either Th1 cells (which are specialized in facilitating cytotoxic responses, e.g., against tumors or grafts) or Th2 cells (which produce a different set of lymphokines important for humoral immune reactions), that, by the way, are mutually suppressive [5,6]. It appears that CD8+ T cells also can acquire Th1-like or Th2-like phenotypes [7]. Moreover, it has long been known that B cells can “switch” to the production of different Ig classes, a choice that is determined by the preponderance of Th1 or Th2 cytokines. It may be important to note that an inappropriate class of response can be pathogenic. As an example, the elicitation of immunoglobulin E (IgE) antibodies (with Th2 help) against pollen is pathogenic and thus will cause allergy, whereas a preponderant IgG response (with Th1 help) against the same antigen will have no (or little) pathological consequence. Similarly, the development of an inappropriate Th1 (instead of Th2) response against organ-specific autoantigen can ha...
Table of contents
- Cover
- Half Title
- Title Page
- Copyright Page
- Dedication Page
- Preface
- Contents
- Contributors
- Part I: Immunomodulation
- Part II: Immunomodulating Agents in Disease
- Part III: Immunomodulating Agents in Therapy
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Immune Modulating Agents by Thomas F. Kresina in PDF and/or ePUB format, as well as other popular books in Medicine & Immunology. We have over 1.5 million books available in our catalogue for you to explore.