Understanding parasite biology and impact is essential when giving advice on parasite control in farm animals. In the first review devoted to parasites of domestic cattle and sheep alone, this book provides in-depth, focused advice which can be tailored to individual farms. It considers the impact of parasites, both as individual species and as co-infections, as well as epidemiological information, monitoring, and diagnostic procedures. Supported throughout by diagrams and photos to aid diagnosis, it also reviews the basis for control measures such as the responsible use of parasiticides, adaptive animal husbandry and other management practices.This book: Focuses on common parasites of domestic sheep and cattle; Places emphasis on understanding host responses and epidemiology so that the impact and seasonality of parasitism can be incorporated into advice and decision making;Highlights the fundamental importance of the individual farm and farmer in assessing endemic parasitism and tailoring control options accordingly;Provides a comprehensive reference listing, including important historical citations, to underpin the content.An important resource for students, veterinarians and researchers of farm animal health, this book maintains a focus on ruminant parasitology in order to deliver evidence-based advice and also context for the application of basic research.

eBook - ePub
Parasites of Cattle and Sheep
A Practical Guide to their Biology and Control
- 392 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Information
Topic
MedicineSubtopic
Veterinary Medicine1 The Origin and Evolution of Parasitism in Domestic Ruminants
Introduction
Parasitism is one of the most successful lifestyles in nature (Poulin and Morand, 2000) and, although it is impossible to know precisely how many parasite species exist (Poulin, 2014; Strona and Fattorini, 2014), it has been estimated that parasitic species considerably outnumber all the free-living species on Earth (Windsor, 1998). There are at least 50% more parasitic helminth species than vertebrate hosts, and among mammals, each individual animal can harbour two cestode, two trematode and four nematode species of parasite over its lifetime (Dobson et al., 2008).
There are several definitions of parasitism, all of which describe an association between one organism (the parasite) and another (the host) in which the parasite derives some benefits, whereas the host gains no advantage and may be harmed. Although there are a few parasites that can switch to a non-parasitic life cycle, for example, the threadworms, Strongyloides spp. and blowflies, which can feed on non-living substrates such as carrion, the vast majority of parasite species are dependent on their hosts for all or part of their life cycle (Smyth, 1962).
Parasites
Parasites can be subdivided into those species that are found inside the host (endoparasites) and those that live in or on the skin (ectoparasites). While there are some exceptions, endoparasites are typically helminths, comprising nematodes (roundworms), trematodes (flatworms) and cestodes (tapeworms); most ectoparasites are arthropods, either insects or arachnids. Strictly speaking, protozoa originally meant early (eukaryotic) life forms; however, in parasitology the word has become synonymous with single-celled organisms. Bacteria (prokaryotes), archaea and viruses are the domain of microbiologists, while fungal diseases of animals, though sometimes included within parasitology (Euzéby et al., 2005), are now usually considered separately.
Evolution of Parasitism
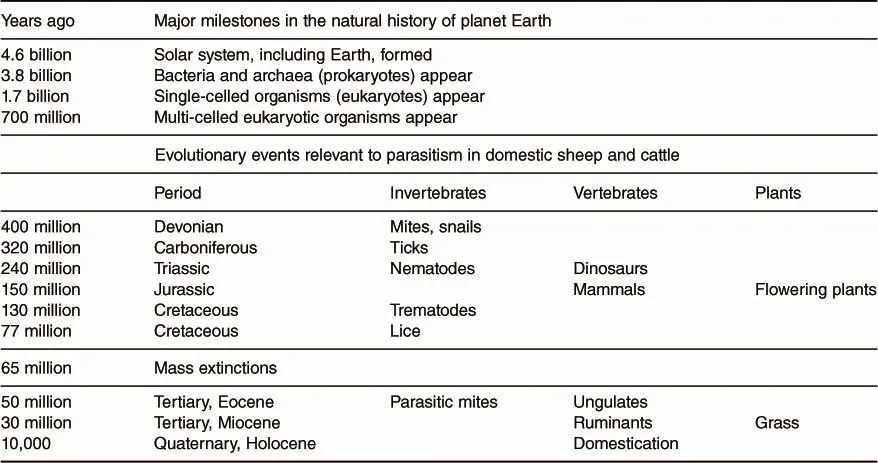
Parasitism has evolved multiple times from free-living invertebrates belonging to diverse phyla that had been present on Earth for millennia (Dorris et al., 1999; Nagler and Haug, 2015). The evidence for the origin and evolution of parasites comes largely from fossils; however, there are obvious limitations of the fossil record for small, soft-bodied organisms that leave little or no direct evidence of their existence. Nonetheless, through the fossil record and adoption of newer molecular biology techniques (Donoghue and Benton, 2007), a more complete picture of the chronology of parasitism is possible, albeit the precise timing of events will inevitably change somewhat as new discoveries are made and new methodologies adopted.
Additional sources of useful information on parasites that transit the gastrointestinal (GI) tract are coprolites, which are fossilized dung and which can contain remnants of parasite eggs. Currently, the earliest fossil evidence for intestinal parasitism in vertebrates stretches back to the Triassic period 240 million years ago (MYA) when ascarid eggs were found in dinosaur coprolites (Poinar, 2015). Younger specimens of dinosaur coprolites from the Cretaceous period (130 MYA) yielded not only nematode eggs and protozoal remains but also trematode eggs, representing an early record of parasitic flatworms (Poinar and Boucot, 2006).
Fossil evidence points to the evolution of both chewing and sucking lice somewhat later than the helminths, probably around 77 MYA, coincident with the initial radiation of mammals (Johnson and Clayton, 2003; Light et al., 2010), though lice also parasitize birds, which diversified earlier. Free-living oribatid mites, which are the intermediate hosts for several species of tapeworms, for example Moniezia spp., have been found in fossils nearly 400 MYA (Arillo et al., 2012), but the obligate parasitic mites of vertebrates have a more recent history with fossil remains found in the Eocene period, ~50 MYA (Walter and Proctor, 2013).
Because of their chitinous exoskeletons, arthropods are preserved as fossils more readily than helminths and protozoa, while another rich source of insect and arachnid remains is amber (Nagler and Haug, 2015), which was formed from plant resins deposited from ~320 MYA. Amber with remains of arthropods currently dates from ~125 MYA, at which time parasitism among invertebrates is evident, for example, a fly parasitized by a Leptus spp. mite (Arillo et al., 2018). Some remarkably well-preserved specimens in amber have shown that ticks parasitized animals 99 MYA (Peñalver et al., 2017), though it has been estimated that ticks may have been present much earlier as parasites of dinosaurs, around 320 MYA (Klompen et al., 1996; Barker et al., 2014).
From this very brief review of some of the evolutionary history of parasitic organisms, it is clear that by the end of the Cretaceous period and the mass extinction that followed ~65 MYA, predecessors of all the main classes of parasite were present in the world’s fauna. Potential intermediate hosts, such as snails, had also evolved from around 400 MYA onwards (Wanninger and Wollesen, 2019). By this time, other important links in the terrestrial food chain were also present, for example, dung beetles (Chin and Gill, 1996) and remnants of grass were found in coprolites from herbivorous dinosaurs in the late Cretaceous period, suggesting the possibility of early grazing mammals at that time (Prasad et al., 2005). Table 1.1 provides approximate evolutionary temporal relationships among the components that eventually led to parasitism in domestic ruminants (Dawkins, 2004; Lloyd, 2009).
Table 1.1. Chronology of evolutionary events relevant to parasitism in domestic sheep and cattle.
The Ruminants
Following the Cretaceous extinctions of multiple species, notably the dinosaurs, and also many invertebrate and plant species (Macleod, 2013), the scene was set for the extensive radiation of flowering plants and mammals from their initial appearance ~200 MYA. The first ungulates (herbivorous, hoofed mammals) to make their presence felt were the perissodactyls (odd-toed ungulates, such as horses, rhinos and tapirs), species of which proliferated from ~55 MYA (Janis, 1976). The artiodactyls (even-toed ungulates, such as camels, pigs, hippos, deer, antelopes, sheep and cattle) evolved later; evidence of the earliest ruminants dates from ~40 MYA and from the late Miocene onwards (~10 MYA), the artiodactyls assumed numerical dominance over the perissodactyls (Janis, 1976).
A characteristic of these herbivores, which is well represented in the fossil record, is the presence of hypsodont teeth; these are large, high-crowned molars with hard enamel ridges that have evolved to grind down plant cell walls to release their contents and to reduce particle size to facilitate bacterial colonization and digestion in the alimentary tract (Janis and Fortelius, 1988). Hypsodont teeth are particularly important in grazing animals as many grass species have a high silicon content, which makes them particularly abrasive. In ruminants, these teeth are central to the efficiency of digestion when fibrous material is regurgitated, re-chewed, crushed and ground and then re-swallowed in the process known as rumination (Hofmann, 1989).
There are around 200 extant or recently extinct species of ruminant (Fig. 1.1), the largest family of which is the Bovidae (bovids), comprising ~137 species of cattle, sheep, goats and antelopes; the next largest family is the Cervidae (deer) with ~47 species (Hernandez Fernandez and Vrba, 2005). The ~150 living ruminant species range in size from <10 kg to >1 t and, although they have several features in common, there is quite a marked variation in their digestive tracts, which reflects adaptation to different diets. Feeding patterns can be categorized as (Hofmann, 1989; Clauss et al., 2010):
• Grazers, feeding predominantly on grass
• Browsers feeding on forbs, leaves and twigs and fruit
• Intermediate feeders, which are opportunist grazers or browsers

Fig. 1.1. Ruminant diversity: wildebeest and springbok.
Domestic cattle and sheep are grazers, while goats are intermediate feeders and their digestive systems are adapted to handle their feed. There are a number of challenges for ruminants living on a plant-based diet, which differ according to their chemical composition. For example:
• Grass and roughages are high in fibre, which has to be broken down by the rumen microflora and fauna, not only in order to digest the cellulose itself but in so doing to release soluble intracellular carbohydrates, proteins and other nutrients. Chewing the cud is required to reduce the particle size of plant material and increase its surface area so that the rumen microorganisms can access the substrate more efficiently.
• Browsers tend to eat young leaves and fruit, which are more easily digestible, particularly the latter, and therefore the breakdown of cellulose is less critical. However, many plants have chemical defences, such as tannins, to deter ...
Table of contents
- Cover
- Half Title
- Title
- Copyright
- Contents
- Preface
- Acknowledgements
- 1 The Origin and Evolution of Parasitism in Domestic Ruminants
- 2 Parasitic Gastroenteritis in Cattle
- 3 Parasitic Gastroenteritis in Sheep: Teladorsagiosis and Trichostrongylosis
- 4 Parasitic Gastritis in Sheep: Haemonchosis; and Parasitic Enteritis in Lambs: Nematodirosis
- 5 Tapeworm Infections
- 6 Parasitic Enteritis: Coccidiosis
- 7 Lungworm Infections in Cattle
- 8 Lungworm Infections in Sheep
- 9 Liver Fluke
- 10 Rumen Fluke
- 11 The Lancet or Lanceolate Fluke
- 12 Obligate Ectoparasites of Cattle: Lice and Mange Mites
- 13 Ectoparasites in Cattle: Flies
- 14 Ectoparasites of Sheep: Sheep Mange Mites
- 15 Sheep Ectoparasites: Insects
- 16 Ticks and Tick-borne Disease
- 17 Grazing Management and Helminth Control on Stock Farms
- 18 Responsible Use of Parasiticides
- 19 Principles and Practical Implementation of Parasite Control on Livestock Farms
- Index
- Cabi
- Back
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Parasites of Cattle and Sheep by Andrew B Forbes in PDF and/or ePUB format, as well as other popular books in Medicine & Veterinary Medicine. We have over 1.5 million books available in our catalogue for you to explore.