
- 450 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Biology of Fishes
About this book
The VitalBook e-book version of Biology of Fishes is only available only in the US and Canada at the present time. To purchase or rent please visit http://store.vitalsource.com/show/978-1-1341-8631-0. The Third Edition of Biology of Fishes is chiefly about fish as remarkably efficient machines for coping with the many problems that life in wat
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Publisher
Taylor & FrancisYear
2008Print ISBN
9781138582965
9781138357716
Edition
3eBook ISBN
9781134186303
1 The Diversity of Fishes
1.1 Introduction
Biologists interested in fishes are very fortunate today not only in having a very wide range of living fish to study, from hagfish to lungfish, but also in being able to examine fishes adapted to every kind of aquatic habitat (and even some that spend most of their lives out of water). In consequence, we can examine fascinatingly different designs for special modes of life, as well as ways used by different fish for solving problems common to them all. Thus, for example, we can look at the remarkable ways in which fish eyes are adapted for seeing in different environments, or the different merits of methods of internal or external fertilization. These kinds of comparisons are often illuminating, and permit insights into the compromises fishes have to make to satisfy the conflicting demands of their lives. To understand these, we need first to have at least a basic idea of what kinds of fishes live today, their structural features, how they are related to each other, and how they arose.
There are sizeable numbers of species in four of the vertebrate classes: Amphibia 4300; Reptilia 6000; Aves 9000; Mammalia 8000. These are all round numbers, but, even so, they are almost certainly unlikely to be added to significantly by the discovery of new species. More likely, extinction (often our fault by habitat reduction) will reduce them. But the fish class “Pisces” is different to the others, on two counts. First, there are more kinds of fish than all other vertebrates added together. So far, over 25 000 different species of fish are known, and around 100 more are described each year, so that the final total may well exceed 30 000. The great majority are bony teleosts, the cartilaginous elasmo-branchs coming a very poor second with just over 800 species. All other fish groups have insignificant numbers of species, although they may be of great zoological interest, and sometimes, as are the sturgeons, of considerable gastronomic and economic importance.
Second, within each class, all the different species should share a common basic morphology. In the class Aves, for example, we see at once that, however modified, ostriches and hummingbirds have a similar basic structure. On structural diversity, the class Aves deserves only to rank as an order of the class Reptilia; separating birds from reptiles is simply a matter of convenience rather than strict taxonomic logic.
In the class Pisces, it does not at first sight seem obvious that, say, sharks and lampreys might reasonably be in the same class. Much emphasis has always been placed (correctly) on the acquisition of jaws as a very significant step in vertebrate evolution, and although it would probably be more logical to put the agnatha in a class of its own, the current “Pisces” is too deeply ingrained to abandon. Indeed, a recent (1997) text suggests that the class Pisces is not monophyletic, that is, derived from a common ancestor, but rather, a mere convenience lumping various unrelated fish groups together! Yet certain features found in all kinds of living fish make it certain that they are all ultimately derived from a common ancestor.
One of the strongest clues to common ancestry lies in the hind brain. Here, in lampreys, in different kinds of bony fish, lungfish, and in some embryo sharks, there are paired large Mauthner neurons found in relation to nerve VIII. They send their large axons down the opposite side of the spinal cord to the cell body, linked to segmental motoneurons, and in many teleosts, command the rapid tail flip escape startle response (Chapter 11, p. 372). Mauthner neurons are not found in all fish, for such a response is not suited to all, and they disappear in adult sharks. They are absent in hagfish, but, although not all would agree, the most recent molecular evidence seems to make it clear that hagfish and lampreys (Figure 1.2) are more closely related to each other than to gnathos-tomes (e.g. Cotton and Page, 2002; Takezaki et al., 2003; Delsuc et al., 2006).
Another clue comes from the pattern of innervation of the myotomal muscles along the body. With the exception of higher teleosts (which have “invented” a new pattern), in all living fishes the rapidly contracting myotomal muscle fibers are innervated at their ends, a pattern shared with urodele amphibia (Chapter 3, p. 71). Another clue from the nervous system comes from the rapidly contracting myotomal muscle fibers in the myotomes, a pattern shared by all fish groups (and urodele amphibia) with the exception of higher teleosts.
With morphological clues of this kind, backed up by an increasing flood of molecular data, we can be confident that all living members of the class Pisces ultimately share a common ancestor.
What was this common ancestor like, and when was it around?
To be recognizable as a chordate, it must be able to show serial muscle blocks or myotomes along its body, and to have the recognizable remains of an axial notochord. Experiments with the little living acraniate amphioxus (Branchiostoma; Figure 1.1), have shown that in animals experimentally “fossilized” (Kear, 1993), the myotomes and notochord can survive for long periods when the animals decay in anoxic conditions, and, indeed, some fossils show both. Fossils of these soft-bodied chordates will certainly be very exceptional; yet there are a few. Among the bizarre fossils of the middle Cambrian Burgess shales in the Rockies, the small Pikaia is an amphioxus-like fossil with V-shaped myotomes, a notochord and, apparently, a bifid head. Pikaia remained on its own for almost a century (and now seems off the main chordate line), but recent finds from the remarkable early Cambrian fossil lägerstetten fields of Chengjiang in China (Shu et al., 1999; Hou et al., 2002) have included various small fish-like chordates (Cathaymyrus, and Myllokunmingia), and more are likely to turn up. They have V-shaped myotomes, with rather more complex gills than amphioxus, and (like amphioxus) lack a bony skeleton. These early fish-like chordates were swimming around some 530 million years ago (mya), and, as a group, their ancestors perhaps arose in the Ediacaran before that. Unsurprisingly, the chordate notochord and myotomal muscle design for oscillatory swimming seems to have been part of the makeup of a number of these early fossils, and not enough of other features can be discerned to enable them to be fitted into even a tentative classification. A recent molecular attempt to dethrone amphioxus from its position as the living prototype ancestor is amusingly discussed (with approval) by Holland (2006)
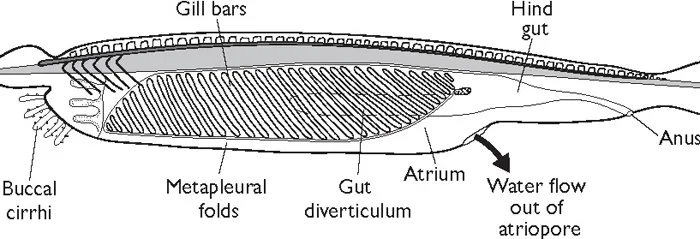
Figure 1.1 The little (5 cm) amphioxus (Branchiostoma lanceolatum) which lives buried in coarse sand, filtering its food on a mucus filter using the current from its ciliated gill bars.
Not very much more is known of a rather different group of early chordates living from the pre-Cambrian to the late Triassic, called the conodonts. Most conodont remains are tooth-like little fossils, built from calcium phosphate, which are known worldwide and are abundant enough to be useful in stratig-raphy. All conodonts used to be lumped together, and still may be in some texts, but the earliest, or protoconodonts, probably have nothing in common with the later conodonts. They seem likely to be related to arrow worms or chaetognaths, and there is no evidence for any chordate affinity.
However, the later conodonts long remained one of the most intriguing puzzles in paleontology, for, until recently, only scattered isolated teeth had ever been found. In 1983 the first body outline associated with conodont teeth was discovered, and, subsequently, other better preserved specimens have been found. Astonishingly, these were flattened impressions of early chordate animals! They were rather fish-like, with notochord, V-shaped myotomes, caudal fins, bone-like material in the “teeth,” and anterior large eyes. Some Carboniferous conodonts had a curious and complex three-dimensional skeletal pattern in the gullet, rather as if the animal had swallowed the wooden parts of a mixed set of folding deckchairs. This conodont apparatus is nothing like anything in any living animal and just how it operated is (unsurprisingly!) arguable. The relationship of conodonts to living forms has yet to be agreed (some texts suggest a relationship with hagfish, on rather tenuous grounds). What these remarkable fossils show us is that there seem to have been several kinds of fish-like animals with notochords, sense organs, and serial myotomes swimming around in the earliest Cambrian and later seas, before clearly recognizable fishes appeared. From one group of these chordates, all living fish are descended.
This not to say that all early kinds of fossil fishes can be fitted into a mono-phyletic scheme with living fishes. Fragments of fossil fishes with a bony skeleton first appeared in the mid-Cambrian some 600–500 mya, and by 500 mya in the Ordovician a variety of these jawless fishes has been found. Naturally enough, systematists have tidily felt the need to link, for example, fossil agnathan fishes with living hagfish and lampreys (Figure 1.2) and some of these fossils may indeed be close to the ancestry of living forms, but, at present, without further information, schemes such as Figure 1.2A are arguable. A recent proposal by two paleontologists is shown in Figure 1.2B. Note that, in this scheme, hagfish and lampreys are not closely linked but are sister groups, and the relationships of fossil groups are cautiously vague!
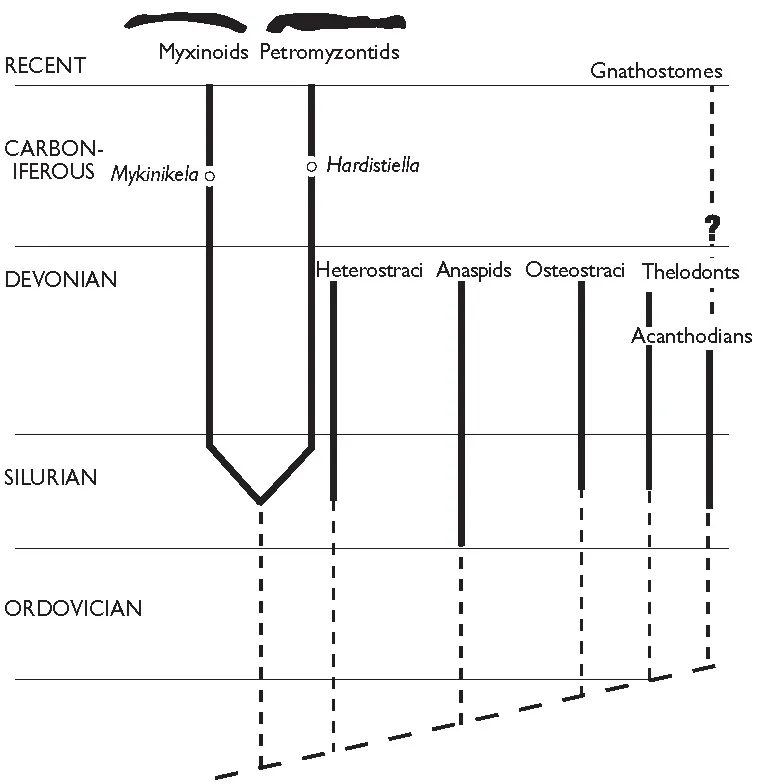
Figure 1.2A One possible arrangement of fossil and recent agnathans, and the origin of the gnathostomes. Modified from Forey and Janvier (1993).
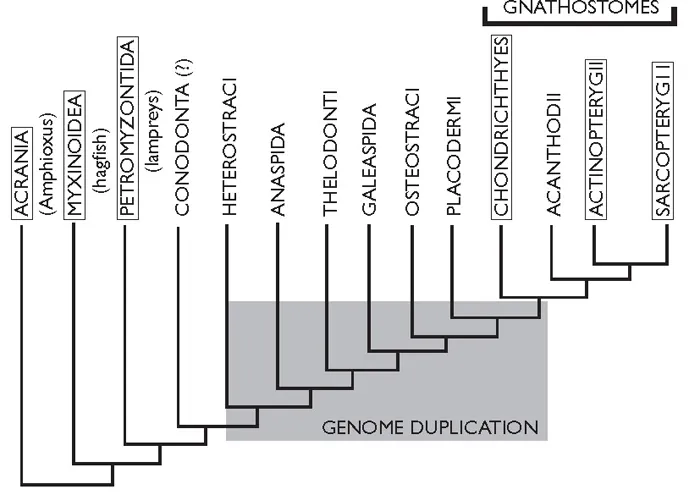
Figure 1.2B Another (more recent) view of relationships between living (boxed) and extinct (unboxed) groups, from tunicates to gnathostomes. Somewhere during the shaded area, gnathostome gene duplication event(s) took place. From Donoghue and Purnell (2006).
1.2 Fish Classification
What can now be called the classical view of the relationships of the major surviving groups of fishes is shown in Figure 1.3. It is based almost entirely on structural features, and was first proposed in the middle of the past century. In its broad outlines, most ichthyologists would have accepted it, although at time of writing there is still disagreement about the correct position of hagfish, and a more modern cladogram would be presented as Figure 1.4.
The names for the different groupings, such as elasmobranchii (plate gills) or coelacanths (hollow spines), may seem rather arcane, but the student should realize that they define the main characters of each. It is well worth looking up these and other names to find out what they mean, as it is easier to remember names that one understands and which contain some information about the fishes, instead of learning them by rote. Although the names seen in Figures 1.2–Figure 1.7 are not found in every text; some of the least fashionable have been retained since the reader may come across them elsewhere. Readers of a less advanced age than the senior author are not, on the whole, recommended to follow the example of the distinguished American ichthyologist David Starr Jordan (President of Stanford University) who (sensibly) refused to learn the names of his students on the grounds that if he did so he would forget the names of fishes.
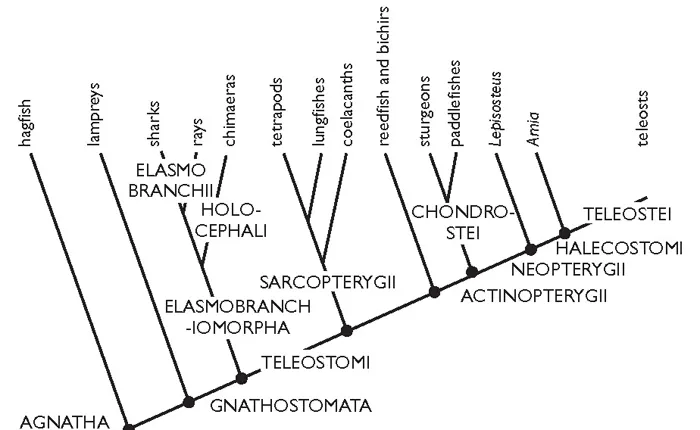
Figure 1.3A The relationships of living fish groups. Mainly after Nelson (2006).

Figure 1.3BCladogram showing relationships between living fish orders. The numbers refer to higher gr...
Table of contents
- Cover Page
- Title Page
- Copyright Page
- Preface
- 1: The Diversity of Fishes
- 2: Fishes and their Habitats
- 3: Swimming
- 4: Buoyancy
- 5: Gas Exchange, Blood, and the Circulatory System
- 6: Osmoregulation and Ion Balance
- 7: Food and Feeding
- 8: Reproduction, and Life Histories
- 9: Endocrine Systems
- 10: Sensory Systems, and Communication
- 11: System The Nervous
- 12: System The Immune
- 13: Cognition Behavior and
- 14: Aquaculture and Fisheries
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Biology of Fishes by Quentin Bone,Richard Moore in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Ecology. We have over 1.5 million books available in our catalogue for you to explore.