Analysis and Application of Analog Electronic Circuits to Biomedical Instrumentation, Second Edition helps biomedical engineers understand the basic analog electronic circuits used for signal conditioning in biomedical instruments. It explains the function and design of signal conditioning systems using analog ICs-the circuits that enable ECG, EEG,
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Before describing and analyzing the electronic circuits, amplifiers, and filters required to condition the signals found in clinical medicine, biomedical research, and physiology, it is appropriate to describe the sources and properties of these signals (i.e., their bandwidths, amplitude distributions, and noisiness). Broadly speaking, biomedical signals can be subdivided into two major classes: (1) Endogenous signals, which arise from natural physiological processes and which are measured within or on the surface of living creatures (examples include electrocardiogram (ECG), electroencephalogram (EEG), respiratory rate, temperature, blood pressure, blood glucose). (2) Exogenous signals that are applied from without (generally noninvasively) to measure internal structures and physiological parameters. These include, but are not limited to, ultrasound (imaging and Doppler), X-rays (CAT scans), and monochromatic light (such as the two wavelengths used in transcutaneous pulse oximeters, excitation of fluorescence from fluorophore-tagged cells and molecules stimulated with blue or near UV light, optical coherence tomography (OCT), laser Doppler velocimetry (LDV) used to measure blood velocity, and the strong magnetic fields used in magnetic resonance imaging (MRI)). For many other examples of exogenous signals refer to Northrop (2002).
In the following section, the properties of endogenous bioelectric signals used in medical diagnosis, care, and research are examined.
1.2 SOURCES OF ENDOGENOUS BIOELECTRIC SIGNALS
The sources of nearly all bioelectric signals are transient changes in the transmembrane potential observed in all living cells. In particular, bioelectric signals arise from the time-varying transmembrane potentials seen in nerve cells (neuron action potentials and generator potentials) and in muscle cells, including the heart and smooth muscles. The electrochemical basis for transmembrane potentials in living cells lies in several phenomena: First, cell membranes are semipermeable. That is, they have different transmembrane conductances and permeabilities for different ions and molecules (e.g., Na+, K+, Ca++, Cl−, glucose, and proteins). Second, cell membranes contain “ion pumps” driven by metabolic energy (e.g., ATP). The ion pumps actively transport ions and molecules across cell membranes against energy barriers set up by the transmembrane potential and/or concentration gradients between the inside and outside of the cell. In the steady state, there is a continual leakage of ions into a cell (e.g., Na+), out of a cell (e.g., K+), and ongoing ion pumping to restore the steady-state concentrations. In the widely studied squid giant axons, the steady-state internal concentrations are as follows: [Na+]i = 50 mM, [K+]i = 400 mM, and [Cl−]i = 52 mM. The steady-state external concentrations (in extracellular fluid) are as follows: [Na+]e = 440 mM, [K+]e = 20 mM, [Cl−]e= 560 mM, and [A−]i = 385 mM (Kandel et al. 1991). [A−] is the equivalent concentration of large, impermeable protein anions in the cytosol. Ion concentration data exist for the neurons and muscles of a variety of invertebrate and vertebrate species (Kandel et al. 1991; Katz 1966; West 1985).
The steady-state transmembrane potential can be modeled by the Goldman–Hodgkin–Katz equation (Guyton 1991):
where T is the Kelvin temperature, R is the MKS gas constant (8.314 J/(mol K)), F is the Faraday number, 96,500 Cb/mol, and PX is the permeability for ion species X.
The resting transmembrane potential of neurons, Vmo, varies with species, neuron type, ionic environment, and temperature. It can range from ca. 60 to 90 mV, inside negative with respect to outside. Muscle cells also have transmembrane potentials, ca. 80 < Vmo < 95 mV, inside negative, for human muscles.
1.3 NERVE ACTION POTENTIALS
Nerve action potentials (APs) are, in general, the result of transient changes in specific ionic conductances and permeabilities induced in the nerve cell membrane electrically or chemically by neurotransmitters. In excitable neuron membranes, an increase in cell membrane sodium permeability leads to a depolarization of the transmembrane potential (i.e., sodium ions flow rapidly into the neuron’s cytoplasm down a concentration gradient and electric field). The inrush of Na+ through the membrane causes the normally negative transmembrane voltage Vm to go positive, which is a depolarization. When the excitable nerve transmembrane voltage reaches a depolarization threshold, which is on the order of a few millivolts more positive than the resting potential, the permeability events that lead to a propagating action potential or nerve spike begin to occur. First, there is a further, “all-or-nothing,” large transient increase in sodium permeability causing a strong transient inrush of Na+ ions. This Na+ inrush causes a large, rapid depolarization so that Vm actually goes positive by 10s of mV, generally in less than a millisecond. Immediately, permeability to K+ ions also increases, but at a slower rate, which causes an outward potassium ion current density,
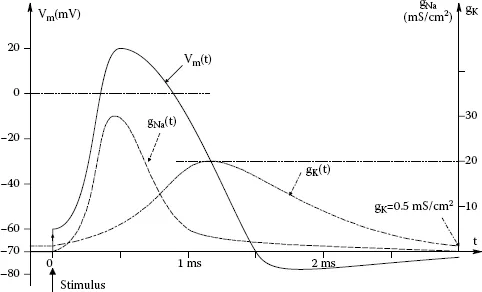
, causing Vm to decrease from its positive peak to its negative resting value after a slight, transient undershoot (hyperpolarization). The total duration of the nerve action potential spike, including recovery to the SS Vm, is on the order of 2.5–3 ms. Once initiated, an action potential propagates down a neuron’s axon at a velocity that depends on a number of physical and chemical factors, including the diameter of the axon. One of the earliest mathematical models for nerve impulse generation was given by Hodgkin and Huxley (1952). The H-H model now appears to be somewhat oversimplified with its description of a single type of potassium channel, but is still valid, and is a useful model to demonstrate the dynamics of nerve impulse generation. Figure 1.1 illustrates the result of a computer simulation of the H-H model using Simnon™ (Northrop 2001). Shown is the transmembrane voltage, Vm(t), and the time-varying conductances for Na+ and K+. Readers interested in pursuing the molecular and ionic details of neurophysiology should consult the texts by Kandel et al. (1991), West (1985), Guyton (1991), and Northrop (2001).
Most neurons in the vertebrate central nervous system (CNS) are too small to directly record their transmembrane potentials with glass micropipette electrodes. However, their action potentials can be recorded over long periods of time with extracellular, metal microelectrodes whose uninsulated tips are in the neuropile within several micrometers of axons or cell bodies. Action potentials from peripheral nerve bundles can be recorded with simple platinum hook electrodes, saline-filled suction electrodes, or saline-wetted wick electrodes coupled to silver/silver chloride electrodes. All extracellular recording techniques suffer from the problem that the electrodes pick up nerve spikes from active, adjacent, or neighboring neurons. This neural background noise is added to the desired unit’s signal and, unfortunately, has the same bandwidth as the desired unit’s spikes. In dissected peripheral nerve fibers, it may be possible to isolate single axons with hook, suction, or wick electrodes, greatly improving the recording signal-to-noise ratio (SNR).
FIGURE 1.1 Results of a Simnon™ simulation of the Hodgkin–Huxley 1952 mathematical model for nerve action potential generation (Northrop, R.B., Introduction to Dynamic Modeling of Neuro-Sensory Systems. CRC Press, Boca Raton, FL, 2001.) Traces shown: vm(t) (transmembrane potential) in mV. gK(t, vm) in mS/cm2. gNa(t, vm) in mS/cm2. Time in ms.
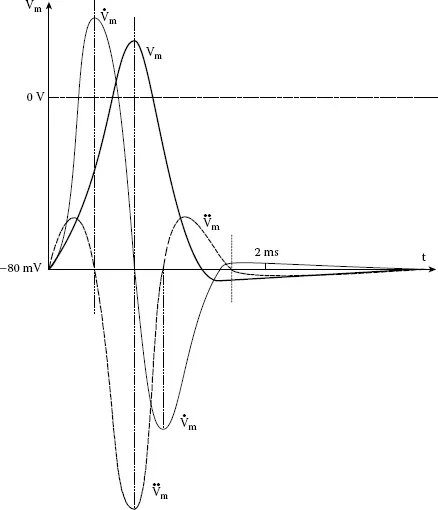
Because the nerve action potential is a traveling wave, it can be shown that an external electrode in close proximity to the outside surface of an axon (the second, (reference), (−) electrode is located remotely from the axon) will respond to the passage of the AP with an electric potential waveform that has in effect, the shape of the second derivative of the transmembrane spike waveform (Plonsey 1969), as shown in Figure 1.2. (The intracellular AP, Vm, is to scale, but its derivatives are not to scale.) The triphasic (second derivative) waveform of the AP recorded at a point near the axon comes from the fact that the AP is traveling along the axon with velocity, v. As the AP approaches the electrode, a weak, net outward
causes a low positive voltage peak. When the AP has moved opposite the electrode, the electrode responds to the strong inward flow of JNa with a large negative voltage peak. Then, as the AP passes the electrode, its potential again goes positive from the net outward
in the recovery phase of the AP.
In the author’s experience, using fine platinum–iridium extracellular microelectrodes (which were glass insulated down to 6–12 μm of their conical tips), we were able to record from single units in insect optic lobes and protocerebrum, and in frog tectum, with major spike amplitudes ranging from −50 to −500 μV. Midband gain for signal conditioning was 104, and signal conditioning bandwidth was typically 100 to 3000 Hz (Northrop and Guignon 1970).
Nerve APs recorded through the neuron membrane (in the cell body, base of dendrites, or axon) using glass micropipette electrodes can be ca. 100 mV or more, peak-to-peak. A capacity-neutralized, electrometer head stage used to couple the high resistance microelectrode generally has a gain of 2 or 3; the second stage may have gain from 5 to 30, so the overall gain can range from 10 to 90. Bandwidth is generally from dc to 3–5 kHz. The direct coupling is required because we are generally interested in the neuron’s resting potential, Vmo, or slow changes in Vm caused by incoming excitatory or inhibitory signals. If we are not interested in Vmo, then it is technically simpler and less noisy to use extracellular microelectrodes and band-pass filtering (e.g., 100 Hz to 3 kHz). More amplifier gain will be required with external electrodes, however.
FIGURE 1.2 A nerve action potential and its first and second time derivatives (derivatives not to scale).
1.4 MUSCLE ACTION POTENTIALS
1.4.1 Introduction
An important bioelectric signal that has diagnostic significance for many neuromuscular diseases is the electromyogram (EMG), which can be recorded from the skin surface with electrodes identical to those used for electrocardiography, although in some cases, the electrodes have smaller areas than those used for ECG (<1 mm2). To record from single motor units (SMUs), or even individual muscle fibers (several of which comprise a SMU), needle electrodes that pierce the skin into the body of a superficial muscle can also be used. (This semi-invasive method obviously requires sterile technique.) EMG recording is used to diagnose some causes of muscle weakness or paralysis, muscle or motor problems such as tremor or twitching, motor nerve damage from injury or osteoarthritis, and pathologies affecting motor end plates (myoneural junctions).
1.4.2 The Origin of EMGs
There are three types of muscle in the body, viz, striated, cardiac, and smooth. Striated muscle in mammals can be further subdivided into fast and slowmuscles (Guyton 1991). Fast muscles are used for fast movements; they include, for example, the two gastrocnemii, laryngeal muscles, the extraocular muscles. Slow muscles are used for postural control against gravity; they include the soleus, abdominal muscles, back muscles, neck muscles, and so on. EMG recording is generally carried out on both slow and fast skeletal muscles. It can also be done on less superficial muscles such as the extraocular muscles that move the eyeballs, the eyelid muscles, and the muscles that work the larynx.
A particular striated muscle is inn...
Table of contents
Cover
Biomedical Engineering Series
Title Page
Copyright Page
Contents
List of Figures
Preface
Author
Chapter 1 Sources and Properties of Biomedical Signals
Chapter 2 Properties and Models of Semiconductor Devices Used in Analog Electronic Systems
Chapter 3 Differential Amplifier
Chapter 4 General Properties of Electronic, Single-Loop Feedback Systems
Chapter 5 Feedback, Frequency Response, and Amplifier Stability
Chapter 6 Operational Amplifiers and Comparators
Chapter 7 Introduction to Analog Active Filters
Chapter 8 Instrumentation and Medical Isolation Amplifiers
Chapter 9 Noise and the Design of Low-Noise Signal Conditioning Systems for Biomedical Applications
Chapter 10 Digital Interfaces
Chapter 11 Modulation and Demodulation of Biomedical Signals
Chapter 12 Power Amplifiers and Their Applications in Biomedicine
Chapter 13 Wireless Patient Monitoring
Chapter 14 RFID Tags, GPS Tags, and Ultrasonic Tags Used in Ecological Research
Chapter 15 Examples of Special Analog Circuits and Systems Used in Biomedical Instrumentation
Appendix
Glossary
Bibliography and Recommended Reading
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Analysis and Application of Analog Electronic Circuits to Biomedical Instrumentation by Robert B. Northrop in PDF and/or ePUB format, as well as other popular books in Technology & Engineering & Biotechnology in Medicine. We have over 1.5 million books available in our catalogue for you to explore.