This respected graduate-level textbook provides comprehensive and accessible coverage of the basic and clinical aspects of the mucosal immune system, addressing the major components of the mucosal barrier ? gastrointestinal, upper and lower respiratory, ocular, and genitourinary mucosal immune systems ? in a highly user-friendly style. The editors of and contributors to the book, all internationally-recognized leaders, present the current principles, concepts, and basic processes involved in mucosal immunology, mucosal diseases, and host defense at mucosal surfaces. Topics discussed include the development and structure of the mucosal immune system and its cellular constituents, host-microbe relationships, infection, mucosal diseases, and vaccines. The second edition has been carefully updated throughout to reflect the latest developments from clinical research and key literature has been fully updated.
eBook - ePub
Principles of Mucosal Immunology
- 596 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Principles of Mucosal Immunology
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
MedicineSubtopic
Clinical MedicinePart One
Development and structure of mucosal tissue
1Overview of the mucosal immune system structure
2Phylogeny of the mucosal immune system
3Immunologic and functional differences among individual compartments of the mucosal immune system
4Secreted effectors of the innate mucosal barrier
1
Overview of the mucosal immune system structure
Reinhard Pabst and Per Brandtzaeg*
Pioneering experiments in the early 1960s showed that the large lymphocytes (lymphoblasts) that enter the bloodstream from the thoracic duct migrate into the intestinal lamina propria and undergo terminal differentiation into plasmablasts and plasma cells. Many of the circulating lymphoblasts express surface IgA (sIgA), and in the gut contain cytoplasmic IgA. These lymphoid cells were thought to be derived mainly from Peyer’s patches (PPs), because transfer studies demonstrated that PPs and the draining mesenteric lymph nodes (MLNs), in contrast to peripheral lymph nodes and the spleen, were enriched precursor sources for IgA-producing plasma cells in the gut mucosa. It was also shown that plasma-cell differentiation occurs during dissemination of the mucosal B cells. Thus, the fraction of cells with cytoplasmic IgA increased from an initial 2% in PPs to 50% in MLNs and 75% in thoracic duct lymph, and finally 90% in the intestinal lamina propria.
These seminal studies led to the introduction of the term “IgA cell cycle,” and subsequent research showed that B cells bearing other sIg classes than IgA, as well as T cells, when activated in PPs, also exhibit gut-seeking properties. It later became evident that different secretory effector sites can receive activated memory/effector B cells from a variety of mucosa-associated lymphoid tissue (MALT).
This work gave rise to the notion that the mucosal immune system is divided into distinct inductive sites and effector sites. The inductive sites are the organized MALT structures together with mucosa-draining lymph nodes, whereas the effector sites are the mucosal epithelia and the underlying lamina propria, which contains stromal cells and associated connective tissue stroma. The mucosae and related exocrine glands harbor by far the largest activated B-cell system of the body, and the major product—J-chain-containing dimeric IgA together with some pentameric IgM—is immediately ready for external transport by the polymeric immunoglobulin receptor (pIgR) across the secretory epithelium and into the mucus layers at mucosal surfaces, to provide antibody-mediated immunity (see Chapter 9).
IMMUNE INDUCTIVE LYMPHOID TISSUE
The MALT concept was introduced to emphasize that solitary organized mucosa-associated lymphoid follicles and larger follicle aggregates have common features and are the origin of T and B cells that traffic to secretory effector sites. This functional distinction is important because while the different tissues can be identified and discriminated by histology, single cell suspensions prepared from mucosal surfaces contain a mixture of cells from small MALT structures that cannot be dissected out, and connective tissue. This is particularly a problem in man, because solitary follicles cannot be seen in resected bowel.
1.1 Mucosa-associated lymphoid tissue is different from lamina propria or glandular stroma
MALT is subdivided according to anatomical regions (Table 1.1), and the cellular content of these lymphoid structures depends on whether the tissue is normal or chronically inflamed, superimposed upon striking age and species differences (Figure 1.1). The PPs in the distal small intestine of humans, rodents, and rabbits are classical MALT structures. PPs are inductive compartments generating conventional (B-2) sIgA-expressing memory/effector B cells and T cells, which, after a journey through lymph and peripheral blood, enter the gut mucosa. This “homing” appears to be antigen independent, but local antigens penetrating into the lamina propria contribute to local retention, proliferation, and differentiation of the extravasated lymphoid cells. Thus, although the lamina propria is considered an effector site, it is clearly important for the expansion and terminal differentiation of T and B cells. Some expansion of mucosal memory/effector T cells can also occur in the mucosal surface epithelium, and there is considerable “cross talk” between these two effector compartments.
| Table 1.1 Different regions of malt and their components | |
| Region | Components |
| GALT (Gut-associated lymphoid tissue) | Peyer’s patches (PPs) and isolated lymphoid follicles constitute the major part of GALT, but also the appendix is included. |
| NALT (Nasopharynx-associated lymphoid tissue) and tonsils | Rodents lack tonsils but do have paired NALT structures dorsally in the floor of the nasal cavity. |
| In humans, tonsils consist of the lymphoid tissue of Waldeyer’s pharyngeal ring, including the adenoids (the unpaired nasopharyngeal tonsil) and the paired palatine tonsils. Scattered isolated lymphoid follicles may also occur in nasal mucosa (NALT). | |
| BALT (Bronchus-associated lymphoid tissue) | Not generally detectable in normal lungs of adult humans. |
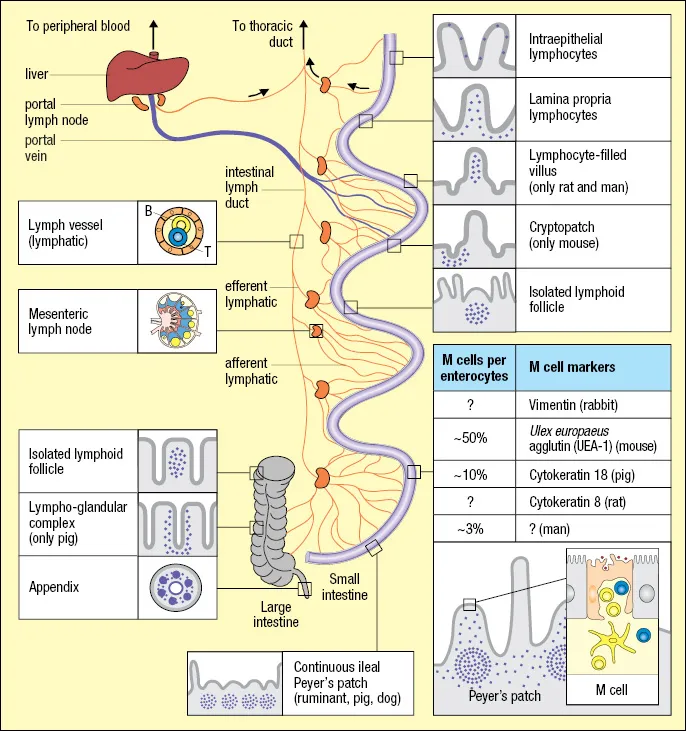
The major component of human MALT comprises the gut-associated lymphoid tissue (GALT), including the PPs, the appendix, and numerous solitary follicles (see Figure 1.1) now termed “solitary isolated lymphoid tissue” (SILT) (Herbrand et al. 2008). Induction of mucosal immune responses can also take place in nasopharynx-associated lymphoid tissue (NALT), the tonsils (pharyngeal, palatine, lingual, pharyngeal), and bronchus-associated lymphoid tissue (BALT) as described later. Moreover, small MALT-like lymphoid aggregates are present in the conjunctiva and are associated with the larynx and various ducts such as those connecting the ocular and nasal compartments.
MALT resembles lymph nodes with B-cell follicles, interfollicular T-cell areas, and a variety of antigen-presenting cells (APCs) but lacks afferent lymphatics and a capsule. MALT therefore samples exogenous antigens directly from the mucosal surfaces through a follicle-associated epithelium (FAE), which contains “membrane” or “microfold” (M) cells (see Chapter 13). These specialized thin epithelial cells do not act as APCs but are effective in the uptake of microorganisms and other particulate antigens; they are also vulnerable “gaps” in the mucosal barrier. Studies in the mouse have, in addition, shown that dendritic cells (DCs) in the dome region of MALT send processes through the FAE to sample gut antigens.
The distinction between mucosal inductive and effector sites is not absolute, as the signals for extravasation and accumulation of naive versus memory/effector B and T cells are different. It is therefore confusing when MALT is used to refer to the mucosal effector compartments (e.g., the lamina propria and the surface epithelium of the gut and its diffusely distributed immune cells, Figure 1.2). This is in conflict with the classical definition of a lymphoid tissue, and so the Society for Mucosal Immunology and the International Union of Immunological Societies have agr...
Table of contents
- Cover
- Half Title
- Title Page
- Copyright Page
- Dedication
- Contents
- Preface
- Contributors
- Part I: Development and structure of mucosal tissue
- Part II: Cellular constituents of mucosal immune systems and their function in mucosal homeostasis
- Part III: Microbial Commensalism
- Part IV: Genitourinary Tract
- Part V: Nose, Airways, Oral Cavity, and Eyes
- Part VI: Infectious Diseases of Mucosal Surfaces
- Part VII: Specific Immune-Mediated Diseases of Mucosal Surfaces
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Principles of Mucosal Immunology by Phillip Smith, Richard Blumberg, Thomas MacDonald, Society for Mucosal Immunology, Phillip Smith,Richard Blumberg,Thomas MacDonald,Society for Mucosal Immunology,Phillip D. Smith,Richard S. Blumberg,Thomas T. MacDonald, Phillip D. Smith, Richard S. Blumberg, Thomas T. MacDonald, Society for Mucosal Immunology in PDF and/or ePUB format, as well as other popular books in Medicine & Clinical Medicine. We have over 1.5 million books available in our catalogue for you to explore.