Originally published in 1977, this title describes the basic structure and function of the brain, as well as the highest cognitive functions, using data from various disciplines to detail ways in which behaviorally relevant functions are mediated by the neural systems. Among the topics discussed are the neurophysiology of emotion, the chemical basis of memory, daily subjective experience and psychopathology, and information representation. A major purpose of this volume was to provide the student not only with a sound foundation in functional neuroscience, but also to equip them with a detailed understanding of how these facts and methods can be applied to clinical problems.

- 382 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Foundations of Cognitive Processes
About this book
Information
1
Basic Neurophysiology
I. Fundamentals of Neurophysiology
This chapter is intended to provide a brief outline of the fundamentals of neurophysiology. More detailed information on this subject can be found in the following references: Brazier (1958); Ochs (1965); Purpura (1959); Quarton, Melnechuk, and Schmitt (1967); and Ruch, Patton, Woodbury, and Towe (1965).
A. Basic Structure
The brain is constituted of three basic compartments: glial cells, the extracellular space, and nerve cells or neurons. Glial cells, consisting primarily of astroglia and Oligodendroglia, form a close and interactive relationship with neurons. They provide metabolic and structural support for the nerve cells. They also appear to serve a reciprocal, “symbiotic” function by exchanging proteins with neurons (Hydén, 1967) and by regulating ionic concentrations in both the intracellular and extracellular space (Karahashi & Goldring, 1966; Nicholls & Kuffler, 1964). The extracellular space and glial cells together provide the structural and functional matrix in which the neurons are embedded.
The extracellular space (sometimes referred to as the intercellular space) is the smallest (15 to 20%) of the compartments. However, through this space flow essential metabolic substances and ionic materials related to the polarization and depolarization of neuronal membranes and related as well to the relatively powerful currents generated by the activity of synchronized masses of neurons. The term “extracellular space” is misleading. Actually, this space is occupied by an extracellular structure. Electronmicrographs reveal an extracellular “fuzz” made of branching mucopolysaccharides and glycosaccharides extending from a protein backbone on the outer surface of neuronal membranes (Fig. 1.1). The complexly branching saccharides contain positively and negatively charged sites that bind extracellular ions. Weak electrical currents, such as those used in measurements of tissue impedance, pass through the extracellular space with very little, if any, penetration of the membranes of neurons or glia (Cole, 1940). Extracellular fluids have a specific resistance of approximately 4 Ω cm”2 as compared with resistances of neurons or neuroglia which are approximately 5000 Ω cm”2 (Coombs, Curtis, & Eccles, 1959; Nicholls & Kuffler, 1964).
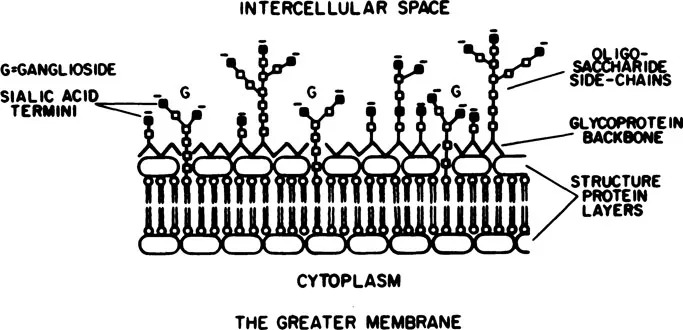
FIG. 1.1 Model of the neuronal membrane illustrating the glycoprotein and polysaccharides that form the extracellular “fuzz.” (From Lehninger, 1968.)
Impedance measurements which reflect conductance within the extracellular channels demonstrate changes during learning, suggesting that structural alterations of the extracellular matrix underlie information storage and retrieval (Adey, Kado, & Didio, 1962; Adey, Kado, Didio, & Schindler, 1963; Adey, Kado, Mcllwain & Walter, 1966b). The greatest concentration of extracellular mucopolysaccharides is around synaptic junctions, the axon hillock, and axonal nodes, three regions which are fundamental in the generation of electrical impulses in neurons (Meyer, 1969). Conformational changes in the extracellular matrix may modify neural excitability by affecting ionic binding affinities at these critical sites. As will be emphasized in later sections, the electroencephalogram (EEG) involves complex current flows within small channels that branch profusely throughout this extracellular matrix.
The third compartment is comprised of neurons. The neuron consists of three functional portions: (a) a cell body or soma which contains genetic material and protein synthesizing mechanisms. The soma also contains specialized receptive membrane structures (at synapses) that allow the neuron to be influenced directly by other neurons. Extending from the soma are (b) dendrites which can be considered as receptive surfaces and (c) an axon which transmits impulses (Fig. 1.2). An axon is usually several times longer than its diameter and in the peripheral nervous system can be as great as one meter in length.
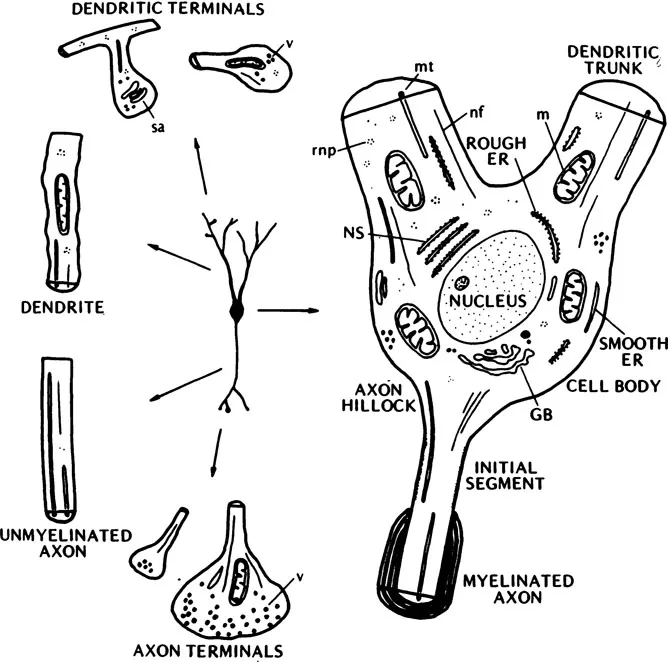
FIG. 1.2 An illustration of a neuron showing the cell body, axon, and dendrites as well as the interneuronal structure. ER, endoplasmic reticulum; GB, Golgi body; NS, Nissl substance; mt, microtubule; nf, neurofilament; rnp, ribonucleic particles; sa, spine apparatus; v, vesicles; m, mitochondria. (From Shepherd, 1974.)
Neurons are electrically excitable. Slow electrical potentials generated in the receptive regions (called synaptic potentials) give rise to impulses (called spikes or action potentials) that are transmitted down the axon. The axon arises from the soma at a specialized region called the axon hillock.
In an electronmicroscope, the soma can be seen to be limited by a double membrane structure which encloses a number of subcellular constituents: endoplasmic reticulum and ribosomes, nucleus and nucleolus, mitochondria, golgi apparati, and neurotubules. The neurotubules form a complex network serving the function of transporting cellular constituents to dendrites and axon terminals. Recent evidence shows that cell bodies strategically located in the brain stem (nucleus coeruleus, nucleus raphe, nucleus substantia nigra) give rise to axons that travel great distances to synapse with the limbic system, the cortex, and the extrapyramidal motor system. Through these axons flow chemical transmitters (norepinephrine, serotonin, dopamine) which play important roles in the regulation of emotion, arousal, and motor-sensory coordination. These transmitters are probably transported through the neurotubules. The function of these chemicals will be discussed in Section III.
The importance of neurotubules has been further emphasized by the discovery that aging is accompanied by a flattening of the dendritic neurotubules (Wis-niewski, Coblentz, & Terry, 1972). Malfunction of the transport mechanisms appears to be responsible, in part, for diminished function of brain cells in the very old.
In addition to the double membrane structure seen in the soma, long axons are enclosed by an insulating cover (myelin sheath) which functions to enable rapid and constant transmission velocities. The myelin sheath is in reality two membranes formed by specialized glial cells (Schwann cells) that wrap around the axon many times to form a multilayered structure (Fig. 1.3). The myelin is interrupted at regular intervals of approximately 0.5 to 1.5 mm along the fiber, by Nodes of Ranvier. The distance between the nodes, the internodal length, vary with axon diameter, age, and species (Hiscoe, 1947; Tasaki, 1953). The myelin sheath is a prominent feature of the long-axoned neurons; that is, those which form the peripheral sensory and motor systems and the long association bundles within the central nervous system (CNS). In the shortest axoned cells only one layer of myelin is present. As a consequence, a myelin sheath is not evident in the electronmicroscope. Over 97% of the cells comprising the CNS (called interneurons or association neurons) have very short axons without myelin sheaths.
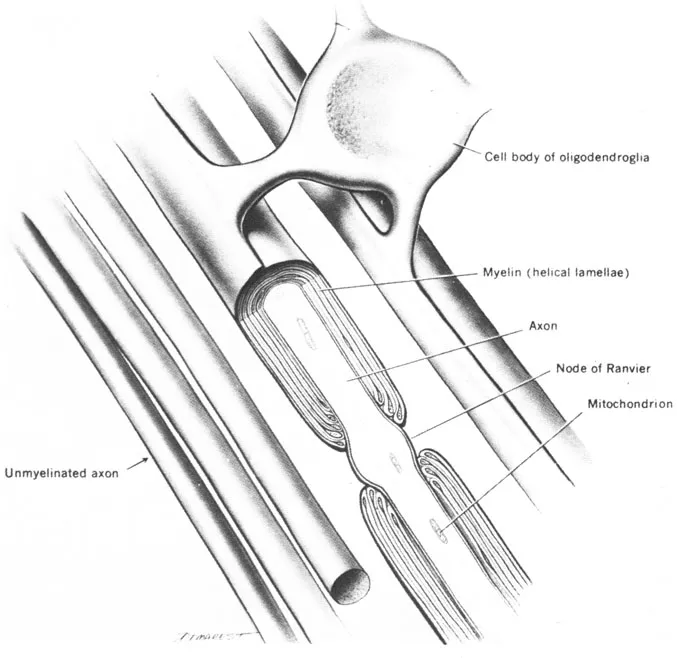
FIG. 1.3 The formation of myelin by Oligodendroglia. (From The Human Nervous System by C. R. Noback & R. J. Demarest. Copyright © 1975 by McGraw-Hill Book Company. Used with permission of McGraw-Hill Book Company.)
B. Action Potential
In the resting state, the inside of the axon is approximately 90 mV more negative than the outside. This potential difference, sometimes referred to as the transmembrane potential or simply the resting membrane potential, is caused by an imbalance in the concentration of ions (particularly sodium and potassium) across the membrane. Figure 1.4 illustrates the various ionic constituents that give rise to the potential difference. The concentration gradients are created by an active metabolic pump mechanism. When a metabolic poison such as Ouabain is administered, the gradients disappear.
There are two forces that act to alter the distribution of the ions across the membrane. One is caused by the individual concentration gradients for each ion, which tend to drive sodium into the cell and potassium and chloride out of the cell. The second is caused by the electromotive force produced by the imbalance of total ions across the membrane, which tends to drive potassium and sodium into the cell and chloride out of the cell. A succinct description of these interactive forces is given by the Nernst equation which expresses the equilibrium potential, that is, the condition where the potential difference across the membrane for a given ion is zero. The Nernst equation is
where V is the equilibrium potential for the ion, R is the gas constant, T is the absolute temperature, Z is the ionic valence, F is the Faraday (which equals 96,500 coulombs), and the quantities in parentheses are the concentration of the ion inside the membrane, (C)i, and outside the membrane, (C)o.
Due to concentration differences, the tendency for CI- ions and K+ ions to diffuse out of the cell is almost counterbalanced by the resting membrane potential. This balanced situation is not present in the case of Na+. Both the concentration gradient and the electrochemical force tend to drive Na+ into the cell.
The resting equilibrium potentials expressed by the Nernst equation are ~ -97 mV for K+ and ~ - 90 mV for CI-. The electromotive force necessary to achieve equilibrium for a resting membrane potential of-90 mV, is -7 mV for K+ and 0 m...
Table of contents
- Cover
- Half Title
- Title Page
- Copyright Page
- Original Title Page
- Original Copyright Page
- Table of Contents
- Preface
- Introduction
- 1. Basic Neurophysiology
- 2. Functional Electrophysiology
- 3. The Genesis of Alpha Rhythms and EEG Synchronizing Mechanisms
- 4. Neurophysiology of Arousal and Attention
- 5. Neurophysiology of Emotion
- 6. Information Representation
- 7. The Neural Representation of Time
- 8. The Chemical Basis of Memory
- 9. The Localization of Function—Where is Memory?
- 10. How Do We Remember?
- 11. Activation of Memories by Electrical Stimulation of the Brain: A Direct Test of Statistical Theory
- 12. Mental Experience
- 13. Daily Subjective Experience and Psychopathology
- References
- Author Index
- Subject Index
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Foundations of Cognitive Processes by Robert W. Thatcher,E. Roy John in PDF and/or ePUB format, as well as other popular books in Psychology & Cognitive Neuroscience & Neuropsychology. We have over 1.5 million books available in our catalogue for you to explore.