Throughout the natural world, organisms have responded to predators, inadequate resources, or inclement conditions by forming ongoing mutually beneficial partnerships--or symbioses--with different species. Symbiosis is the foundation for major evolutionary events, such as the emergence of eukaryotes and plant eating among vertebrates, and is also a crucial factor in shaping many ecological communities. The Symbiotic Habit provides an accessible and authoritative introduction to symbiosis, describing how symbioses are established, function, and persist in evolutionary and ecological time.
Angela Douglas explains the evolutionary origins and development of symbiosis, and illustrates the principles of symbiosis using a variety of examples of symbiotic relationships as well as nonsymbiotic ones, such as parasitic or fleeting mutualistic associations. Although the reciprocal exchange of benefit is the key feature of symbioses, the benefits are often costly to provide, causing conflict among the partners. Douglas shows how these conflicts can be managed by a single controlling organism that may selectively reward cooperative partners, control partner transmission, and employ recognition mechanisms that discriminate between beneficial and potentially harmful or ineffective partners.
The Symbiotic Habit reveals the broad uniformity of symbiotic process across many different symbioses among organisms with diverse evolutionary histories, and demonstrates how symbioses can be used to manage ecosystems, enhance food production, and promote human health.

- 232 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
The Symbiotic Habit
About this book
Information
Publisher
Princeton University PressYear
2021Print ISBN
9780691113425
9780691113418
eBook ISBN
9781400835430
CHAPTER 1
The Significance of Symbiosis
INDIVIDUALS OF DIFFERENT SPECIES form persistent associations from which they all benefit. These relationships are symbioses. The core purpose of this book is to assess the biological significance of symbioses and to investigate the processes by which symbioses are formed and persist in both evolutionary and ecological time. Symbioses are biologically important because they are widespread and dominate the biota of many habitats. I address this aspect of the biological significance of symbiosis later in this chapter. For the present, I suggest that any doubting readers should “look out of the window” and list every organism they can see. I can guarantee that most, probably all, organisms on the list are a product of symbiosis. The prevalence of symbioses is not, however, the only reason why symbioses should be important to biologists. An additional reason is that symbiosis challenges two widely accepted tenets of biology: the universality of descent with modification in evolution, and the primacy of antagonism in interactions among organisms. I will start by explaining these challenges.
1.1 SYMBIOSIS AS A SOURCE OF NOVEL TRAITS
The core expectations of evolution by descent with modification are that morphological, physiological, and other traits of an organism are derived from traits in the ancestors of the organism, and that changes in these traits can be described by multiple, small steps with each intermediate condition viable. Most traits can be explained in this way, but there is unambiguous evidence that some traits of great evolutionary and ecological importance have been gained laterally from different, often phylogenetically distant, taxa. Some laterally acquired traits are novel for the recipient organism and they can be evolutionary innovations, i.e., “newly acquired structures or properties which permit the assumption of a new function” (Mayr 1960, p. 351)].
There are two routes for the lateral acquisition of traits: symbiosis and horizontal gene transfer. Entire organisms are acquired by symbiosis and so the traits gained can be genetically, biochemically, and even behaviorally more complex than those obtained by horizontal transfer of isolated genes. Two very different types of symbiosis illustrate this point. The eukaryotic cells that acquired the cyanobacterial ancestor of plastids gained the capacity for oxygenic photosynthesis as a single package, including many correctly expressed genes, the integrated molecular and cellular machinery for the assembly of multicomponent photosystems in photosynthetic membranes, and the enzymatic machinery for carbon fixation. Similarly, an Acacia tree that is protected from herbivores by a resident colony of ants has acquired a morphologically and behaviorally complex defense capable of responding appropriately to herbivore attacks of variable magnitude and type.
The appreciation that descent with modification is an inadequate evolutionary explanation does not mean that symbioses contradict current understanding of evolutionary processes. Symbioses are subject to natural selection and, contrary to some claims (e.g., Ryan 2003), they have no discernible dynamic independent of natural selection.
1.2 SYMBIOSIS AS A TYPE OF BIOLOGICAL ALLIANCE
Interactions among organisms are routinely portrayed as principally antagonistic, dominated by competition, predation, and parasitism. The history of life has even been described as “a four billion year war” (Marjerus et al. 1996). This perspective is not wrong, but it is incomplete. Organisms have repeatedly responded to antagonists (predators, competitors, etc.) and abiotic stresses such as low nutrient availability by forming alliances, i.e., interactions with other organisms, resulting in enhanced fitness and ecological success of all the participants. As with alliances among people, political parties, and nation states, the persistence of many biological alliances depends on the continued presence of the antagonist, and the benefit gained from the alliance can vary with the identity of the participants and environmental conditions. I return to this issue in section 1.3.
Most alliances are founded on reciprocity, that it is advantageous to help another organism only if the favor is returned. In a two-organism system, reciprocity requires that each of the organisms places a higher value on what it receives (benefit) than what it gives (cost) (figure 1-1a). For example, the relationship between mycorrhizal fungi and plant roots is underpinned by the transfer of photosynthetic sugars from plant to fungus, and of phosphate in the reverse direction. Photosynthetic carbon is cheap for the plant to produce but a critical resource for the fungus, which cannot utilize the polymeric carbon sources in soil. Inorganic phosphate is relatively immobile in soils, and is acquired more readily by the fine, branching fungal hyphae than by the relatively massive plant roots with short, nonbranching root hairs. The one situation where reciprocity as depicted in figure 1-1a does not apply is between closely related organisms. Here, kin selection is important: genotypes that help relatives (i.e., individuals with many genes in common) increase in frequency and are at a selective advantage over genotypes that do not help.
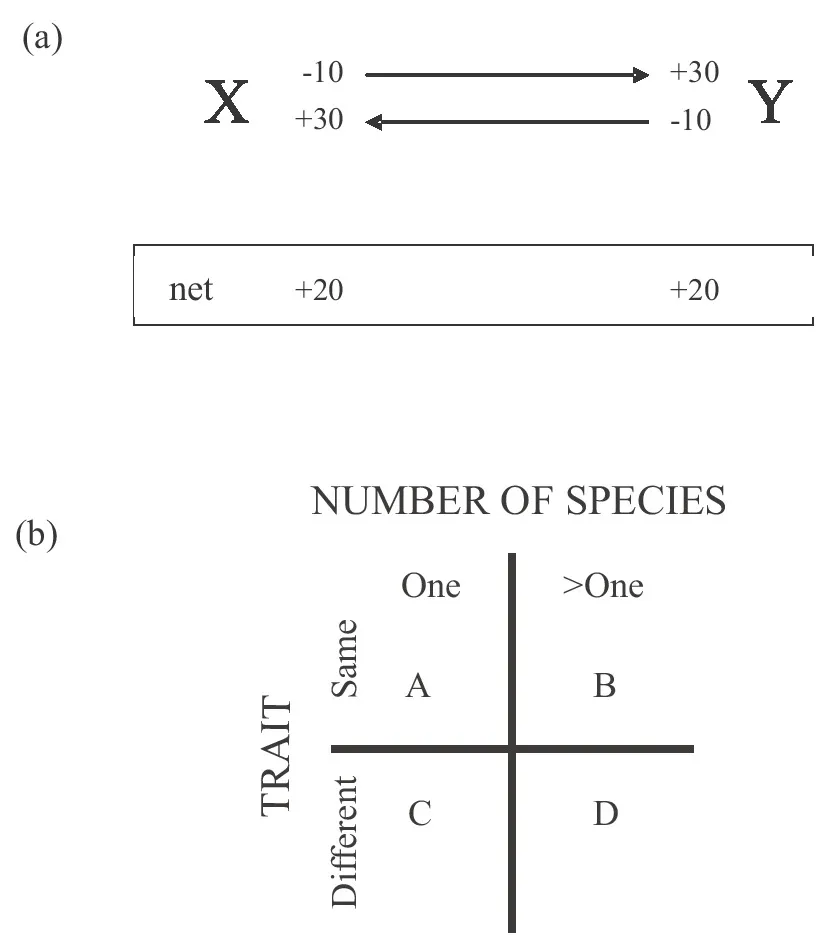
Figure 1-1 Biological alliances. (a) Most alliances are underpinned by reciprocity between the two participating organisms, X and Y. Each organism provides a service at cost of 10 arbitrary units and receives a benefit of 30 units, yielding net benefit of 20 units. (b) Alliances are classified according to the number of species and traits involved. Examples of each type of alliance include (A) roosting bats huddling together, sharing their uniform trait of heat production; (B) mixed-species flocks of passerine birds foraging for food in winter; (C) bartering of goods between humans; and (D) consortia of multiple microbial species that, through their complementary metabolic capabilities, degrade otherwise recalcitrant organic compounds. [Figure 1-1a modified from figure 1 of Douglas (2008)]
Many organisms in alliances display cooperative traits, i.e., traits that are advantageous to another organism (the recipient) and that have evolved because of their beneficial effect on the recipient. The Acacia trees introduced in section 1.1 display cooperative traits that benefit their resident ants: swollen, hollow thorns which provide domatia (nest sites) for the ants, and extrafloral nectar and highly nutritious antbodies on which the ants feed (figure 1-2). In return, various behavioral traits of the ants are advantageous to the plant, including patroling the plant to protect against herbivores and removing fungal spores at the breakpoint of the ant-bodies to prevent fungal infection of the plant. As a contrary example, the food in the gut of an animal infected by a tapeworm is not a cooperative trait because, although the tapeworm benefits from the food, animals have not evolved the habit of eating for the benefit of tapeworms.

Figure 1-2 Adaptations of Acacia sphaerocephala for symbiosis with protective ants of the genus Pseudomyrmex. (a) Hollow thorns that provide domatia (nest sites) for ants. (b) Extrafloral nectary at base of leaf. (c) Lipid-rich ant-body at tip of leaflet. [Reproduced from figure 4-18 of E. O. Wilson (1971) The Insect Societies. Harvard University Press]
Alliances can be classified according to whether they involve one or multiple species displaying the same or different traits, and this two-way classification generates four categories (figure 1-1b). The focus of this book is category D in figure 1-1b, alliances between different species with different traits. These alliances are also known as mutualisms, which are formally defined as relationships from which all participants derive benefit. In this book, I consider symbioses as a type of mutualism, specifically mutualisms in which the participants are in persistent contact.
This brings me to the most frustrating difficulty in the field of symbiosis—the lack of a single universally accepted definition. Disagreement over definitions has led to disputes about which relationships are symbioses and, consequently, a lack of consensus about the common features of symbiotic systems. Two alternative definitions of symbiosis, neither fully satisfactory, have dominated the literature for many decades: “symbiosis as any association” and “symbiosis as a persistent mutualism.” Here, I digress briefly from the core topic of this chapter, the significance of symbioses, to address the thorny problem of definitions.
1.3 DEFINITIONS OF SYMBIOSIS
1.3.1 Symbiosis As Any Association
The term symbiosis was coined originally by Anton de Bary in 1879 to mean any association between different species, with the implication that the organisms are in persistent contact but that the relationship need not be advantageous to all the participants. De Bary explicitly included pathogenic and parasitic associations as examples of symbioses. Many symbiosis researchers use this definition and, without doubt, some colleagues steeped in the symbiosis literature will have objected to the opening two sentences of this book.
One key advantage of the definition of de Bary is that it promotes a broad context for research into symbioses. It acts as a reminder that it is important to investigate both the costs and the benefits to an organism of entering into a symbiosis (see figure 1-1a); and it is reasonable to expect some of the processes underlying relationships that are classified as mutualistic and antagonistic to be similar. For example, just as the persistence of certain antagonistic interactions depends on one organism failing to recognize the antagonist as a foreign organism, so some organisms may be accepted into symbioses because they fail to trigger the defense systems of their partner and not because they are positively recognized as mutualists.
Nevertheless, the definition of de Bary has two serious shortcomings. First and very importantly, the definition is not accepted by most general biologists or nonbiologists today, and so fails to communicate effectively. Most people do not describe the current malaria pandemic or the potato blight that caused the Irish famine of the 1840s as examples of symbiosis. Second, there are few principles generally applicable to symbioses, as defined by de Bary, but inapplicable to other biological systems. As a result, the “symbiosis as any association” definition is something of a catch-all category. Although this definition does promote further enquiry and insight into symbioses, any insights obtained are unlikely to be common to all symbioses defined in this way.
1.3.2 Symbiosis As a Persistent Mutualism
The definition of symbiosis widely accepted among both general biologists and the lexicographers who prepare English dictionaries is an association between different species from which all participating organisms benefit. I subscribe to this definition even though it is not without difficulties.
The “symbiosis as a persistent mutualism” definition requires a formal assessment of the benefit derived by the organisms in the association. The standard approach to identify benefit is to compare an organism’s performance (survival, growth, reproductive output, etc.) in the presence and absence of its partner. If the organism performs better with the partner, it benefits from the relationship and if it performs better in isolation, then it is harmed by the association. Although the methodology appears straightforward, it is unsuitable for many associations.
There are two types of problem. First, for some associations, there are formidable practical difficulties. Consider the deep-sea symbioses, such as the chemosynthetic bacteria in the tissues of pogonophoran worms at hydrothermal vents and the luminescent bacteria in the lure of deep-sea angler fish. It is difficult to envisage how bacteria-free pogonophorans and angler fish could be generated experimentally and how the performance of the bacteria-free individuals could be monitored reliably in habitats so inaccessible to humans.
The second and more fundamental problem is the variability of real associations, such that benefit is not a fixed trait of some relationships but varies, ...
Table of contents
- Cover Page
- Title Page
- Copyright Page
- Contents
- Preface
- Chapter 1: The Significance of Symbiosis
- Chapter 2: Evolutionary Origins and Fates of Organisms in Symbiosis
- Chapter 3: Conflict and Conflict Resolution
- Chapter 4: Choosing and Chosen: Establishment and Persistence of Symbioses
- Chapter 5: The Success of Symbiosis
- Chapter 6: Perspectives
- References
- Index
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Symbiotic Habit by Angela E. Douglas in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Ecology. We have over 1.5 million books available in our catalogue for you to explore.