Genome-editing methods are becoming routine tools for molecular and cell biologists. Such tools include ZFNs, CRISPR, megaTALs and TALENs. These tools are revolutionizing the creation of precisely manipulated genomes to modify the characteristics of organisms or cells. Additionally, gene drives have altered the way we understand inheritance laws. They give us the ability to have total control of the inheritance of traits of choice and importance. This succinct volume summarizes the history, principles and applications – as well as the advantages and disadvantages – of each of these tools and various kinds of gene drives. The book is part of a program to produce books helpful to students and faculties of science at colleges and universities. This volume in the Pocket Guides to Biomedical Sciences series will help demystify these technologies. The book fills the gap between established conventional methods and the novel and exciting newly introduced tools of genome editing and gene drives. It will help young scientists understand the emerging genome-editing tools and gene drives, thereby promoting related research and adoption.
Key Features
Extensively reviews the current genome-editing tools and gene drives
Clarifies the targeting mechanisms and specificity of genome-editing tools
Details many different types of natural and synthetic gene drives
Highlights concerns with gene drives and genome-editing tools
Related Titles
Brown, T. A. Genomes 4 (ISBN 978-0-8153-4508-4)
Samuelsson, T. The Human Genome in Health and Disease: A Story in Four Letters (ISBN 978-0-8153-4591-6)
Soh, J., et al. Genome Annotation (ISBN 978-1-4398-4117-4)
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
The interaction of the environment and the genotype of an organism determine its characteristics. A complete set of an organism’s genetic material is called its genome. The genome is composed of RNA and DNA (Ahmed et al., 2019). Microbes, animals, and plants also typically have either RNA or DNA or both as their genome. All the genetic information for the growth and development of the organism is contained in the genome (The Royal Society, 2016). A long time before the field of genetics was established, humans genetically modified plants through breeding and selection. Without knowledge of genes, mutagenesis, or gene editing, people of old influenced the genetic makeup of plants and animals by selecting for traits that are conducive to food production and culling those that had unfavorable traits (Wang et al., 1999).
Genome editing tools are tools that can be used to edit the genome of any organism by manipulation of the specific gene loci to gain genome modifications, such as insertions, deletions, or point mutations (Baker, 2012). Conventional methods that have been used to alter genomes or genetic material have been largely based on the use of chemicals and radiation. However, imprecise changes to the genetic material are made using these methods, making it difficult to predict results from the modification events. The advent of recombinant DNA technology in the 1970s gave scientists the ability to make more precise changes to genetic material by inserting or deleting genes or bases. Nevertheless, the process was still relatively imprecise and consequently made inaccurate changes to the genetic material.
In the olden days, genetic studies relied on the discovery and analysis of spontaneous mutations. In the mid-twentieth century, Muller (Muller, 1927) and Auerbach (Auerbach et al., 1947) demonstrated that the rate of mutagenesis could be enhanced with radiation or chemical treatment. Later on, methods that relied on transposon insertions that could be induced in some organisms were used, but these procedures, like radiation and chemical mutagenesis, for example, produced changes at random sites in the genomes of organisms. The first targeted genomic changes were produced in yeast and mice in the 1970s and 1980s. This gene targeting depended on the process of homologous recombination, which was remarkably precise but very inefficient in its outcome, for example, in mouse cells. Recovery of the desired products required a powerful selection and thorough characterization. Because of the low frequency and the absence of culturable embryonic stem cells in mammals other than mice, this resulted in gene targeting not being readily adaptable to other species (Thomas et al., 1986). Traditionally, a poor understanding of genomes contributed to the ineffectiveness of gene editing using these tools. The situation, however, changed with the improvement of technology and subsequent sequencing of genomes of various organisms. It is noteworthy that techniques for gene editing that can make more precise and intentional changes to genes and genomes were later developed and are in use (Robb, 2019; Qi, 2017). These techniques can accurately edit, remove, or insert bases or genes in genetic material or genomes (Urnov, 2018). All the gene-editing technologies utilize DNA-cutting enzymes called nucleases. They also have a DNA-targeting mechanism that guides the nuclease to a specific location on the DNA that has to be cut (Zhang et al., 2018). These mechanisms usually involve proteins that make it easy for gene-editing tools to manipulate specific genes in certain areas, as opposed to random gene changes that were made by prior tools (Malzahn et al., 2017). These gene-editing tools are capable of modifying one gene at a time. They typically modify (edit), delete, or insert bases. More advanced gene-editing tools such as those based on the clustered regularly interspaced short palindromic repeats (CRISPR) are capable of editing multiple genes simultaneously. Editing of the genome is referred to as genome editing (Winterberg et al., 2019). These tools were developed in the last 10 years and are still being perfected, although they have high gene-editing efficiencies. Their efficiency is also attributed to the fact that genome sequences for most species are now easily assembled and available. These are used in targeting and guiding the endonucleases (Koch, 2016). Zinc finger nucleases (ZFNs), megaTALs, and transcription activator-like effector nucleases (TALENs) recognize protein and DNA, while CRISPR recognizes RNA and DNA. These facilitate the editing of genes and altering pathways giving scientists the ability to micro-edit DNA codes and mRNA fate via posttranscriptional modifications (Wright et al., 2018). Currently, we have three powerful classes of nucleases that can be programmed to make double-strand breaks (DSBs) at essentially any desired target: ZFNs, TALENs, and CRISPR-Cas. However, CRISPR-Cas now dominates in research laboratories around the world; the other two are also still in use for research and various agricultural and medical research. All of these nucleases arose from investigations into natural biological processes and not from intentions to find genome editing reagents (Carroll, 2014). CRISPR-Cas9 is the most common technology being applied now because it has become cheaper, faster, more accurate, and more efficient than other existing genome editing methods. The Cas9 endonuclease is a system made up of four parts that include two small RNA molecules named CRISPR RNA (crRNA) and trans-activating CRISPR RNA (tracrRNA) (Barrangou, 2014).
Principle
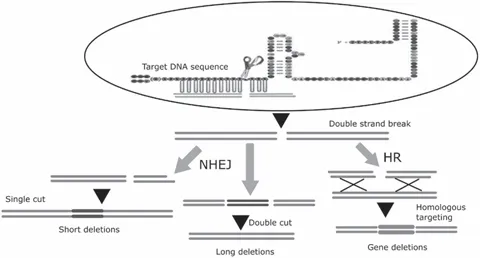
The foundation of the development of targeted genome editing is based on DNA repair mechanisms after its damage and the consequent structural changes in the DNA. A combination of regulatory proteins and sequence-identifiable programmable nucleases makes site-specific genetic and epigenetic regulations possible (Guha et al., 2017). In the last decade, a blooming of targeted genome editing tools and applications have been witnessed in the research community. Programmable DNA nucleases such as ZFNs, TALENs, and the clustered regularly interspaced short palindromic repeats-Cas9 system (CRISPR-Cas9) possess long recognition sites and they can cut DNA in a very specific manner. These DNA nucleases facilitate targeted genetic alterations by enhancing the DNA mutation rate via the introduction of double-strand breaks (DSB) (Source: Roy et al., 2018) at a predetermined genomic site. Figure 1.1 shows DNA editing using tools such as CRISPR, ZFN, and TALENs.
Figure 1.1 Gene (genome) editing of DNA using tools such as ZFN, TALEN, or CRISPR-Cas9 techniques.
A designer nuclease (ZFN, TALEN, or CRISPR-Cas9) cleaves a DNA molecule at a target DNA site to generate a DSB. The DSB can be repaired with one of two endogenous DNA repair mechanisms. These are the nonhomologous end joining (NHEJ) and the homology-directed repair (HDR). The two ends of the DSB are joined together in the NHEJ pathway and ligated without a homologous repair template, which often inserts or deletes nucleotides (indels) to cause gene disruption (knockout). The HDR pathway requires an exogenous DNA template to be provided along with a site-specific genome editing nuclease to repair the DSB, thereby triggering the knock-in of the desired DNA sequence into the genome of an embryo or animal cells (Chandrasegaran and Carroll, 2016).
Compared to conventional homologous recombination-based gene targeting, DNA nucleases, which are also referred to as genome editors (GEs), can increase the targeting rate by around 10,000- to 100,000-fold. The successful application of different genome editors has been shown in a range of different organisms, for example in insects, amphibians, plants, nematodes, and several mammalian species, including human cells and embryos (Gurdon and Melton, 2008). Gene-editing technologies that use programmable nucleases include ZFNs, TALENs, and CRISPR. The CRISPR nucleases are also called RNA-guided engineered nucleases (RGENs). The differences among these are that ZFNs are hybrids between a DNA cleavage domain from a bacterial protein and sets of zinc fingers that were originally identified in sequence-specific eukaryotic transcription factors. TALENs employ the same bacterial cleavage domain but link it to DNA recognition modules from transcription factors produced by plant pathogenic bacteria. CRISPR-Cas is a prokaryotic system of acquired immunity to invading DNA or RNA. ZFNs were a result of the first eukaryotic sequence-specific transcription factor to be characterized, which was found to have zinc-binding repeats in its DNA-binding domain. Related sequences from other transcription factors were shown to be peptide modules (Pavletich and Pabo, 1991). Changing a few residues in a single zinc finger altered its DNA recognition specificity, and fingers could be devised to recognize many different DNA triplets. In the case of TALENs, some plant pathogenic bacteria secrete into host cells proteins that bind to and regulate the activity of host genes to promote the infection. For ZFNs and TALENs, some bacterial restriction enzymes cut DNA a few base pairs away from their recognition sites, and this is because they have physically separable binding and cleavage domains. CRISPR-Cas began with the discovery of a cluster of odd, short repeats in a bacterial genome. Between those CRISPR sequences are short sequences that were shown to match viral genomes (Barrangou et al., 2007). Some CRISPR-associated (Cas) proteins encoded adjacent to the repeat clusters mediate capture of these viral...
Table of contents
Cover
Half Title
Series
Title
Copyright
Dedication
Contents
Preface
Acknowledgments
Authors
Abbreviations
1 Genome Editing
2 Zinc Finger Nucleases
3 TALENs
4 MegaTALs
5 CRISPR
6 Comparison of ZFNs, TALENs, CRISPR, and MegaTALs
7 Gene Drives
8 Supernumerary B Chromosomes
9 Suppression Drives
10 Maternal-Effect Dominant Embryonic Arrest (Medea) System
11 Heritable Microorganisms: Wolbachia
12 CRISPR Gene Drives
13 The Killer-Rescue System
Glossary
References
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Genome Editing Tools and Gene Drives by Reagan Mudziwapasi,Ringisai Chekera,Clophas Zibusiso Ncube,Irvonnie Shoko,Berlinda Ncube,Thandanani Moyo,Jeffrey Godfrey Chimbo,Jemethious Dube,Farai Faustos Mashiri,Moira Amanda Mubani,Duncan Maruta,Charity Chimbo,Mpumuzi Masuku,Ryman Shoko,Rutendo Patricia Nyamusamba,Fortune Ntengwa Jomane in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biotechnology in Medicine. We have over 1.5 million books available in our catalogue for you to explore.