Definitive yet concise, Food Allergy, by Drs. John M. James, Wesley Burks, and Philippe Eigenmann, provides expert guidance for efficient diagnosis and effective management of these increasingly prevalent conditions. The consistent, practical format, with a wealth of case studies, clinical pearls and pitfalls, full-color photos and illustrations, diagrams, and more make this an ideal quick reference tool for both allergy clinicians and primary care physicians.- Quickly reference essential topics thanks to a templated, focused format that includes a wealth of full-color photos and illustrations, diagrams, case studies, and more.- Benefit from the knowledge, experience, and global perspective of leading international authors.- Deliver the best outcomes by incorporating clinical pearls from experts in the field into your practice.- Stay current with timely topics including our latest understanding of non-IgE-mediated food allergies; cross-reactions; future therapies; natural history and prevention; and a review of unproven diagnostic and therapeutic techniques.

- 368 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
MedicineSubtopic
ImmunologyChapter 1 Overview of Mucosal Immunity and Development of Oral Tolerance
Key Concepts

Introduction
The mucosa is the principal site for the immune system’s interaction with the outside environment. Unlike the skin, which is characterized by many layers of stratified epithelium, the intestinal mucosa is lined with a single layer of columnar epithelium. Almost two tons of food travel past this thin barrier each year. More than one trillion bacteria representing about 500 distinct species live in contact with it. The vast majority of these bacteria are non-pathogenic commensals, but pathogens lurk in this diverse antigenic stew, and even the commensal bacteria have the potential to cause harm if not kept in check. The mucosal immune system performs the essential job of policing this boundary and distinguishing friend from foe.
Not only must the mucosal immune system determine the local response to an antigen, but, as the primary site of antigenic contact for the body, it also plays a central role in directing the systemic response to antigens. Oral tolerance – the modulation of the immune response to orally administered antigens – is a fundamental task of the mucosal immune system. In general, as befits the ratio of benign to pathogenic antigens it encounters, the default response of the mucosal immune system is tolerance. The tendency to tolerize to fed antigen can even be used to overcome already developed systemic sensitization, something known and exploited long before the specific cells comprising the immune system were identified. Yet, despite the general bias toward tolerance, the mucosal immune system is capable of producing protective responses to pathogens. This response is controlled by recognition of inherent characteristics of the antigen, or contextual cues such as tissue damage. In general, the immune system is remarkably skilled at responding properly to the antigens it encounters. Failures, albeit uncommon, can be very serious. Food allergy is a prime example of the failure of oral tolerance.
How the mucosal immune system determines when to sound the alarm and when to remain silent is the focus of this chapter. In it, we examine the normal response to food proteins, how that response can go awry, and the factors that tip the balance.
Structure and function
The primary role of the GI tract is to absorb food and liquid and eliminate waste. To achieve this goal, the surface of the tract is both enormous (100 m2) and extremely thin. The lumen of the intestinal tract provides a hospitable environment for bacteria that help break down foods into absorbable nutrients. However, the thinness of the barrier between external and internal creates a grave danger. It is not just nutrients, but toxins, pathogenic bacteria, viruses and parasites that are kept out by a single cell layer only. Breaks in this thin barrier create a risk of systemic infection. The complex task of protecting this border involves both non-specific and highly targeted techniques.
Chemical defenses
Protection begins with chemical and physical measures that keep some of the potentially harmful antigens (both food and microbial) from contact with the mucosal immune system and thus from generating an inflammatory response. Although the intestinal lumen is one of the most microbiologically dense environments in the world, bacteria and large antigens are actually maintained at some distance from the epithelial cells that line the GI tract. This is accomplished by a rich glycocalyx mucin layer (the mucus), which is produced by specialized intestinal epithelial cells. Antimicrobial peptides are caught in the mucous layer in a concentration gradient that provides a zone of relative sterility immediately proximal to the epithelial layer. In mouse models, deficiency of either the mucins or the antimicrobial peptides results in chronic inflammation. In humans, mutations causing abnormal production of the antimicrobial peptides are associated with the autoimmune syndrome Crohn’s disease.1,2 Whether dysfunction in the mucous layer or antimicrobial peptides play a role in the development of food allergy is an area yet to be explored.
What is known is that the enzymatic degradation of food proteins is a first line of protection against allergic sensitization, and that defects in digestion of food antigens contribute to allergy. Many food proteins never get a chance to cause the systemic immune responses characteristic of allergy because they are labile and are denatured by the acidic contents of the stomach. Allergens tend to be proteins that are resistant to this degradation, and thus capable of reaching immune cells to cause sensitization and reaction. For example, β-lactoglobulin and Ara h2, some of the relevant allergens for milk and peanut allergy, respectively, are not denatured by the conditions of the GI tract. Other potential allergens, such as the birch homologs found in many fruits, are easily broken down: although they can induce oral symptoms in cross-reactive individuals, they do not typically initiate sensitization by themselves. Several studies have lent evidence to the importance of the normal enzymatic processes in preventing allergy by showing that antacids impair oral tolerance in both animals and humans. Further, in mice, encapsulation of potentially allergenic foods facilitated allergy by allowing intact allergen to be present in the small intestine.3
The fact that most proteins are broken down by acid and enzymes may help explain why most foods tend not to be allergens, but it does not explain why allergy to stable proteins remains relatively rare. Peanut, for example, contains several proteins that are not degraded, yet only about 1% of the US population is allergic to it, despite near universal exposure. Clearly, other factors come into play after the digestive processes of the stomach.
Trafficking of antigen across the epithelium
Proteins that are not degraded by enzymatic processes can come into contact with the immune system in a number of ways. Transport across the epithelium is both active and passive, occurring both in the spaces between the cells and across them (Fig. 1.1).
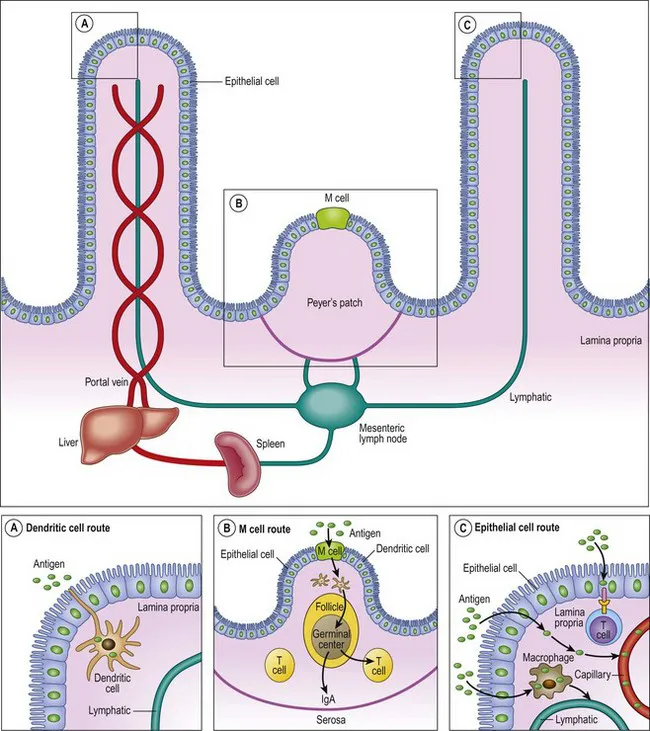
Figure 1.1 Antigen sampling in the gut. (A) Dendritic cells sample antigen directly by extending processes into the lumen. (B) Antigen taken up by M cells travels to the underlying Peyer’s patches. (C) Antigen can cross the epithelium for transport to antigen-presenting cells, T cells, or into the lymphatic circulation.
Reproduced with permission from: Chehade M, Mayer L. Oral tolerance and its relation to food hypersensitivities. J Allergy Clin Immunol 2005; 115: 3–12.
The high-volume route for fluid is via the paracellular spaces, and the overall permeability of the mucosa is regulated by tight junctions that seal the space between epithelial cells. The leakiness of these junctions is subject to a variety of factors, including cytokines, medications and nutritional status. Permeability varies along the GI tract, and even within a short area, as the pores of the villi allow passage of larger solutes than those of the crypt.2,4 Cytokines associated with both autoimmune and allergic disease disrupt barrier function and increase permeability.5 Children with food allergy have been shown to have increased intestinal permeability, both at a time when they are regularly consuming the relevant allergen and after a long period of avoidance.6,7 Other evidence for the importance of barrier function in allergy is the high rate of new sensitization in people taking the anti-rejection medicine tacrolimus, which causes mucosal barrier dysfunction. Although tacrolimus has other effects on the immune system, the high rate of new food allergies after solid organ transplantation is thought to be due its effects on mucosal integrity.5
In addition to the paracellular route, several alternative transport systems actively carry proteins, electrolytes, fatty acids and sugars across cells. Specialized modified epithelial cells called M (or microfold) cells act as non-professional antigen-presenting cells. These cells stud the follicle-associated epithelium overlying specialized collections of immune cells called Peyer’s patches. They express receptors that recognize microbial patterns and aid in the endocytosis and transfer of antigen to the basal surface of the epithelium. This is especially important for bacteria, but may also be relevant for food allergens.4
Other non-specialized columnar epithelial cells form vesicle-like structures that allow transport of dietary proteins across cells. The formation of these vesicle-like structures seems to be dependent on MHC class II binding, but transocytosis can also occur via binding of antigen to IgA, IgE, and IgG. Transport via IgE may be especially important in the acute allergic response and in the amplification of allergy.4 In contrast, secretory IgA, which accounts for the majority of the immunoglobulin produced by the body, complexes with antigen and facilitates transport across the epithelium to antigen-presenting cells, with a tolerogenic outcome.
A final method of antigen transport...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Preface
- List of contributors
- Acknowledgments
- Chapter 1: Overview of Mucosal Immunity and Development of Oral Tolerance
- Chapter 2: Food Antigens
- Chapter 3: The Epidemiology of Food Allergy
- Chapter 4: Clinical Overview of Adverse Reactions to Foods
- Chapter 5: Atopic Dermatitis and Food Allergy
- Chapter 6: Food-induced Urticaria and Angioedema
- Chapter 7: Pollen–Food Syndrome
- Chapter 8: The Respiratory Tract and Food Allergy
- Chapter 9: Food-induced Anaphylaxis and Food Associated Exercise-induced Anaphylaxis
- Chapter 10: Eosinophilic Gastroenteropathies (Eosinophilic Esophagitis, Eosinophilic Gastroenteritis and Eosinophilic Colitis)
- Chapter 11: Food Protein-Induced Enterocolitis Syndrome, Food Protein-Induced Enteropathy, Proctocolitis, and Infantile Colic
- Chapter 12: Approach to the Clinical Diagnosis of Food Allergy
- Chapter 13: In Vivo and In Vitro Diagnostic Methods in the Evaluation of Food Allergy
- Chapter 14: Oral Food Challenge Procedures
- Chapter 15: Management of Food Allergy and Development of an Anaphylaxis Treatment Plan
- Chapter 16: Patient Education and Empowerment
- Chapter 17: Future Therapies for Food Allergies
- Chapter 18: Natural History and Prevention of Food Allergy
- Chapter 19: Diets and Nutrition: Cross-reacting Food Allergens
- Chapter 20: Diagnostic and Therapeutic Dilemmas: Adverse Reactions to Food Additives, Pharmacologic Food Reactions, Psychological Considerations Related to Food Ingestion
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Food Allergy E-Book by John M James,A Wesley Burks,Philippe Eigenmann in PDF and/or ePUB format, as well as other popular books in Medicine & Immunology. We have over 1.5 million books available in our catalogue for you to explore.