First published in 1961, this book provides a contemporary definition of race, the distinction between geographical, local and micro-races, as well as consideration of the major evolutionary mechanisms of race formation in man.
Author Professor Stanley M. Garn was and remains a pivotal figure in the history of biological interpretations of race. He considered racial classification based on physical traits to be imprecise, and believed physical traits to be independent of each other, making classification by the assumption that a population shares certain traits incorrect. He also argued that racial classifications based on physical type seemingly elevated some physical traits to a racial status, but glossed over others, and concluded that racial classifications based on physical type can always be compartmentalized into smaller populations which share more physical traits in common.
Thus, here in his book Human Races, he used three gradations of racial classification which were increasingly more specific in scope: geographical, local and micro.
"Human Races is an attempt to describe what race is, and the mechanisms of racial differentiation in man. It will, I hope, help to dispel the antiquated notions of three "original" races, of the persistence of racial types, and of the role of undirected chance in bringing about racial differences. In their stead, I trust will emerge the contemporary picture of man's genetic response to local selective factors, the constantly changing nature of the natural populations we call races."—Author's Preface

- 118 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Human Races
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Subtopic
Ancient HistoryIndex
HistoryIV — BLOOD GROUPS AND RACE
FOR MANY centuries the possibility of transfusing blood has appealed to surgeons. Great numbers of men died from loss of blood following accidents or battle. Patients expired during and following operations, patients who could have been saved by blood transfusion. But until 1900 transfusions were impractical: too often they ended in shock and death. Somehow, blood just didn’t mix.
Then, about fifty years ago, Moss and Jansky discovered the existence of four different blood types which they named types I, II, III and IV. By determining the blood “type” of the patient, and transfusing compatible blood, blood transfusions became a fortunate practicality. And, as a result of the extensive work on blood-typing, during and since World War I, a tremendous amount has been learned about blood types and their relation to local and geographical races.
THE ABO SYSTEM
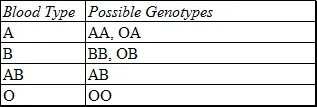
The original four types of Moss and Jansky have since been renamed A, B, AB and O. These blood types are simply inherited with both A and B dominant over O. Genetically, type O blood is OO, while the phenotypes A and B may be either AA or OA, and BB or OB respectively.
Of the three blood types (or factors) in the A-B-O system, O is the most common the world over. Among some American Indian groups, over 90% of the people are of blood group O, with gene frequencies for O as high as 0.99. In much of Europe, the incidence of O is from 35% to 40%, and the frequency of the gene r is between 0.6 and 0.7. Among Chinese, Japanese and Asiatic Indians and in many African populations, O may be as low as 30% with a gene frequency of as little as 0.5. However, even within a particular geographical race there are marked differences in the incidence of O. By way of example, O has been reported in 97% of Utes, but in only 23% of Blackfeet Indians. In Europe there is an increasing incidence of blood type O from south to north, a trend that is repeated even within the British Isles.
Blood group A, the next most frequent blood type the world around, similarly evidences its own pattern of distribution. It is rare (under 5%) in some Amerindian populations, yet extremely common (over 75%) in others; both are world extremes, and point to the wide divergences possible between local races. By way of comparison, the phenotype frequency of A is about 45% in England (with a gene frequency of 0.25), and the same among Americans of North-west European descent.
Blood group B, the least common of the three types in the A-B-O system, has the most interesting world distribution. It is completely absent in most North and South American Indians, rarely over 2% in others, and this may be due to admixture. Blood group B is less common than A in Europe, with a phenotype frequency of 9-25%, averaging about 15%. However, B which increases in frequency to 22% in Ukrainians and 25% in Egyptians, reaches maxima of 35-37% In China, Java, Bengal and the Siberian north. B is an Asiatic and African blood type much more than it is European and it is not at all Amerindian.
Comparing Sumatra, Java and the Philippines with Polynesians, there is a great divergence in A-O-B blood groups. The Asiatic-Malaysian areas are high in B, whereas Polynesia is low in B (under 3%). Clearly Polynesians cannot be derived from any recent mixture with Malaysian people. In similar fashion, the low to zero incidence of blood group B in aboriginal America precludes major recent contact with Asiatic Mongoloids whose frequency of B ranges from 20 to 40% in different populations.
However, blood type A is divisible into two subtypes, A1 and A2; of the two A1 is far more common than A2. The incidence of A2 varies from zero (in Amerindians and Australians) to 10-15% in much of Europe. American Indians agree with Asiatic Mongoloids in the virtual absence of A2, as is true of Polynesia and Australia as well. In fact, A2 is practically limited to Europe and Africa.
Broadly, the A-B-O blood system can be summarized by noting (1) the near absence of B and absence of A2 in the Americas, (2) the low to moderate frequencies of B and increasing frequency of A2 in Europe, and (3) the high incidence of B and relative rarity of A2 in Asia. But the Australian aborigines seem practically Amerindian, as do the Polynesians in their low incidence of B and absence of A2. The A-B-O system, taken alone would suggest a separate origin for them, whereas it groups Asia, India and Africa.
THE MNS-U SYSTEM
Next in order of discovery to the A-B-0 system, and therefore in the amount of information we now have, is the MNS-U system. For years it was the M-N system, then S was added, and more recently U (sometimes written Su).
As with A and B, M and N are inherited without dominance, A person may be M, N or MN, corresponding to the genotypes MM, NN and MN. For much of the world, the frequencies of M and N are about equal, and rarely is either M or N entirely missing. Thus, in England the gene frequencies for M and N are close to 0.53 and 0.47 and in Japan they approximate 0.56 and 0.44 respectively.
But M is peculiarly high among the American Indians, and in many Amerindian tribes there is little or no N. (Low values for N have also been reported for the Near East). In contrast to the situation in America, Australia is the virtual homeland of N, and M—that is MM individuals are entirely absent in some aboriginal hordes. Throughout the Pacific, in Papua, Fiji and Hawaii blood type N is similarly predominant over M. Whereas these peoples of the Pacific are similar to Amerindians in the low incidence of B and of the subtype A2, the high N frequencies of Pacific people set them off completely and preclude recent major contact with America, as their absence of B does with Asia.
Since the extremes of M and N are found in populations formerly explained on the basis of “admixture” it is notable that no combination of Asiatics could yield the low N values common in America, and no Caucasoid-Negroid combinations could yield the nearly M-free peoples of Australia. Either the “original” three races never existed, or subsequent evolution has so altered their genetic makeup in particular localities as to make proof of the three-race hypothesis quite impossible.
Still, two additions to the M-N system must be made. One involves the rare gene S, discovered in 1947, which involves a mutation alternately from M or from N. A person may be MS, MNS or NS. The S gene, discovered in England, (where it is quite common) has since been shown to be absent in Australian aborigines, but present in natives of New Guinea. Tentatively, therefore, one may suggest either closer affinities between Papuans and Europeans, or an absence of ties between the Australian and European geographical race. Perhaps MS and NS might be called M2 and N2, in which case we could say that both M2 and N2 are fairly common in Europe, but absent among the aboriginal Australians.
The second addition to the MNS system involves the gene U, sometimes written as Su. Among Europeans everyone is U-positive. In Negroes, or more specifically the American Colored, about 1% are U-negative as shown below. This polymorphism leads to complications, both in transfusions and in childbirth. Transfusing U-positive blood into a U-negative Negro can be dangerous. While segregated blood stores would not be the answer (most Negroes are U-positive as are nearly all Europeans) the necessity to consider race in planning transfusions becomes all the more apparent. Further divergences in Negro blood groups will be mentioned later in this chapter.
U-NEGATIVE PHENOTYPE AND GENE FREQUENCIES
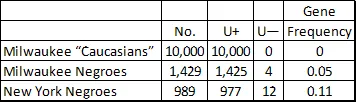
From Greenwalt, Sasaki, Sanger and Race (1956) and Sampson and Thomas (1959).
RHESUS AND RACE
By now, most educated individuals have heard about the Rhesus factor and Rh incompatibility. They know that, especially in later pregnancies, a rhesus-positive fetus can be damaged by antibodies produced by the Rh-negative mother.
Actually, there is a long series of rhesus genes, R1, R2, R0, rʹ, r variously written as Rʹ, Rʺ, etc. Practically, R1, R2, rʹ and others are dominant over the rhesus negative gene, which will therefore be written as r;{6} the rhesus-negative individual is a homozygous rr.
Among Europeans in whom the rhesus-negative gene was first discovered, the proportion of Rh negative individuals ranges from about 12% through an average of approximately 15% in England and the United States, to nearly 30% in Basques.
Elsewhere in the world, rhesus-negative blood is rare, uncommon or even totally abse...
Table of contents
- Title page
- TABLE OF CONTENTS
- PREFACE
- ACKNOWLEDGMENTS
- LIST OF ILLUSTRATIONS
- I - THE NATURE OF RACE
- II - GEOGRAPHICAL, LOCAL AND MICRO-RACES
- III - RACE DIFFERENCES
- IV - BLOOD GROUPS AND RACE
- V - NATURAL SELECTION AND RACE
- VI - ABNORMAL HEMOGLOBINS, MALARIA AND RACE
- VII - RACE AND DISEASE
- VIII - RACE AND GENETIC DRIFT
- IX - RACE MIXTURE
- X - RACE, BEHAVIOR AND INTELLIGENCE
- XI - A TAXONOMY FOR MAN
- REQUEST FROM THE PUBLISHER
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Human Races by Prof. Stanley M. Garn in PDF and/or ePUB format, as well as other popular books in History & Ancient History. We have over 1.5 million books available in our catalogue for you to explore.