
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Pragmatism and the Search for Coherence in Neuroscience
About this book
We have known for over a thousand years that the brain underlies behavioral expression, but effective scientific study of the brain is only very recent. Two things converge in this book: a great respect for neuroscience and its many variations, and a sense of investigation and inquiry demythologized. Think of it as foraging for coherence.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
PhilosophySubtopic
Mind & Body in Philosophy1
Foraging, Learning, and Knowing
We come ready to investigate, to search for coherence and meaning. Embedded in the process of satisfying our basic wants and needs are a lot of problem-solving skills – something I like to call “cephalic capabilities.” I use the word cephalic, an older word not much used in the neurosciences, because of its link to the brain in the context of the body. Our brains are not “a brain in a vat” – that is, a brain detached from action and divorced from the contours of adaptation; rather, a brain is part of a body in space and time that is the rare limiting factor for behavior and experience. This chapter deals with some of the foraging skills our cephalic capabilities allow.
We do not come into the world as a blank slate. Rather, diverse forms of cognitive capabilities permeate neuronal expression and structure at the very onset of life (Carey, 2009). These capabilities include the ability to think about numbers, food, space, time, detecting others and their beliefs, and so on. We also start out with a certain cephalic plasticity. Regions of the brain are modified by experience. While other animals have some plasticity, in humans it is vast: look at the diverse languages we can learn, the many forms of music we can express, the different inventions that we can generate, all of which expand our sense for seeing, hearing, and knowing.
Cephalic systems traverse the whole of sensory and information-gathering systems with which we sample the world and update our orientations. There is no separation of a mind from a body, but there are diverse forms of information sampling of the environment, both internal for what is to be sustained in viable ways and sampling external events for what is occurring.
This capability has been called many things, including “Descartes’ Error” (by the neurologist Damasio, 1994). For some, there is a separation of bodily sensibility from larger scale rational assessment, but many thinkers never made this short-sighted mistake (e.g. Dewey, 1925; Merleau-Ponty 1968). The issue in cephalic functioning (and its evolution by adding on diverse forms of information processing and of sampling and appraising diverse terrains) is its adaptive value and applications.
In this chapter, I begin with a discussion of organized action, inferences about objects, and inquiry and learning in a context of foraging for coherence. Problem solving is a continuous function across the natural and cultural landscape as we look into our evolutionary past, neural systems, and present circumstances (Dewey, 1910).
Codified habits Codified habits are well-organized forms of action. Coherence requires continual appraisal amid codified habits, which was something Charles Sanders Peirce thought of metaphorically as “frozen cognition.” It is a misleading metaphor, perhaps, but it does undercut a mind/body distinction, suggesting information processing that is adaptive and functional for the discernment of predictive discomfort and disappointment (Peirce, 1889). Codified habits are the stuff of everyday action: all the many ways we function in the world. Normal functioning – for example, getting through doors, recognizing faces, deciding to cross at the crosswalk – are all codified habits of behavioral responses.
We also now know that regions of the brain, such as the basal ganglia, underlie codified structural habits (Graybiel, 1998). They are as diverse as the motor possibilities in our species. The systems are finite, but creative capabilities are inherently in cephalic systems, embedded in the “life world” (Schutz, 1932) of foraging for coherence. Noam Chomsky (1965) describes the many forms of linguistic expression available from a finite set of phonemes by the novel combination of events. Novel forms of syntactical expression also take root in diverse motor expression, orchestrated by the basal ganglia.
Syntax does not exist in a vacuum, however. It provides a context for adaptation, sampling, expanding for coherence, foraging, making sense, and problem solving. In a world that matters, survival, long-term reproduction, shorter-term social contact, and comfort are all at stake. Social comfort is no panacea for us. We survive because of our relationships. We were helpless at first, and we still need others to get through our entire lives.
But being alone is also a nontrivial part of who we are. Our capacity to master things on our own and to enjoy doing it is almost as essential to us as is social interaction. Jean-Jacques Rousseau, in Reveries of a Solitary Walker (1776–1778), reveals the connection between self- reflection and human discovery in ten solo strolls in and around Paris and Geneva, moving from descriptions of plants to political philosophy. The man who extolled the sociability of humanity also understood our need for private cogitation.
Within the solitary or the social context, perception is not divorced from framework (i.e. ways of understanding and sampling events). Frameworks are embedded in the reliable and the practical and tied to the predictable. The brain, understood in this context, has a distributed network of perceptual systems (Rolls and Treves, 1998), integrating sensory and more conceptual information as it samples adaptive terrains.
Jerzy Konorski (1948; 1967), the great Polish behavioral neuroscientist, co-discovered with B. F.Skinner (1938) what Konorski called “type II conditioning.” This is what Skinner called operant conditioning; Konorski, working in Pavlov’s laboratory in St Petersburg, showed data to Pavlov (who at first resisted it) revealing another form of cephalic conditioning which was both operant and instrumental (Todes, 2014).
Konorski was oriented to the brain and to understanding the brain. He wrote integrative books similar to the works of Sherrington, spelling out his perceptual theory of cephalic function. Of course, when one considers the diverse forms of visceral information and other kinds of sensory and motor information that pervade cephalic functioning, Konorski was on the right track about the pervasive features of the perceptual – or, as I would put it, the merging of the conceptual and the sensory/motor (Prinz, 2002).
Konorski was seeking to integrate visceral perception and operant behaviors – that is, action and perception combined. An important next step was broadening the learning to what matters, to adaptation in cephalic systems as the prediction of reward, the prediction of things that matter, and events to be avoided.
During the period of the middle to late 1950s, Neil Miller at Yale University began a series of studies that he linked to expanding associations toward predictive relationships (1957, 1959). After all, it is prediction of events that matters in the context of foraging for coherence and adapting to circumstances, both in what is present at hand and in anticipation of what is to come.
Foraging for coherence and sampling environmental stimuli of significance is tied to the fact that, within traditional learning, associations are embedded within probability of occurrences. Learning, in part, takes place when expectations and coherent forms of behavioral adaptation are compromised or called into question (Rescorla and Wagner, 1972).
Within learning theory (Rescorla and Wagner, 1972), prediction is coupled with expectations and their breakdown. When expectations are thwarted, a broad array of learning occurs through new problem-solving and search principles. This is close in scope to Peirce’s (1878) view of inquiry and the development of new solutions to problems. Of course, inquiry is more than this. Anthony Dickinson (1980) demonstrated the link between causal inference and prediction in a wide array of animals. Peirce’s view of inquiry and learning was prescient because the variants of this view would capture learning theory through what became known as the Rescorla-Wagner (1972) equation:
Δ V = α β (λ – V)
The Rescorla-Wagner model depicts the associative strengths of stimuli and how discrepancies from expectations are resolved. An association, and thereby learning, occurs by the strength of the predictions that are being developed. The model then is not simply a mathematical approach to neural science, but also an incorporation of a cognitive point of view. In the equation, V represents the current associative strength of the stimulus, while λ shows the maximum associative strength of the primary motivating event.
The salience of conditioned and unconditioned stimuli is represented by α and β, respectively.
The predictability of the primary motivating event is shown in the (λ – V) term. When the current and maximum associative strengths of the stimulus are equal, the conditioned stimulus fully predicts the reinforcer. However, when the term is positive (λ is greater than V), the associative strength increases and the conditioned stimulus does not fully predict the reinforcer – there is room for learning to occur. With increased associative strength, learning occurs, and in fact, it occurs only when the conditioned stimulus does not entirely predict the reinforcer. In contrast, a negative (λ – V) term occurs when there is a loss of associative strength and the predicted reinforcer has failed (extinction).
General informational search and discrepancy mechanisms, such as the one outlined above, are core features underlying general inquiry, while we “tame chance” (Hacking, 1964) by making such events predictive. As John Dewey (1925) understood it, we search for the stable amid the precarious; we search for the predictive.
Now, learning is certainly much more than the breakdown of the expectation of coherence. It is, however, one part of it, something that the polymath C. S. Peirce, perhaps the first experimental psychologist in America and at the heart of the rise of statistical reasoning in science, suggested in a number of his papers. Peirce emphasized an experimental spirit that pervades his work, along with a logical acumen in the context of hypothesis formation and foraging for coherence.
In one paper, “The Fixation of Belief,” Peirce highlights the context of hypothesis formation and habit generation when expectations are thwarted, when the anticipatory behaviors have lost credence, and when the expected outcomes are undermined. From there comes a further foraging for coherence and propagating actions that are consistent with newly formed hypotheses and expectations.
In his early work, Peirce, like others who overemphasized a variant of drive reduction conception of hypothesis formation (e.g. Freud, 1924; Tinbergen, 1951; Hull, 1943), did not specify enough that learning also happens in cephalic states not linked to failed expectations. Peirce (1878) was aware of this, nevertheless, and his other works emphasize this aspect of exploratory learning.
The science of exploration takes place within a diverse array of cephalic systems designed to facilitate social contact and social meaning (Dunbar, 1995). Making contact is a vital resource in our evolutionary ascent, and it reflects the expansion of diverse forms of cortical and neocortical functions. Stable groups are a core feature of our species, and they are directly related to the encephalization of neural tissue (Dunbar and Shultz, 2007).
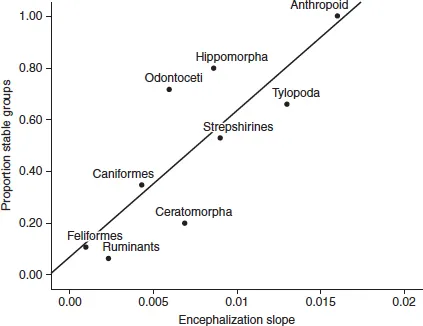
Figure 1.1 Stable groups and encephalization
Source: Adapted from Shultz and Dunbar, 2010.
Science is hypothesis testing amid foraging for coherence, from Aristotle’s taxonomizing through Francis Bacon to the present. Neuroscience is firmly rooted within that domain. But science is also the outgrowth of basic forms of adaptation (Dewey, 1910), expanded and codified into practice, into a “pragmatic realism” (Dunbar, 1992, 2007).
Of course, science also involves a whole array of concepts that are not quite natural (e.g. for Dunbar it is Newton’s action at a distance), but our common sense goes only so far. Yet there is no edifice of certainty; we look for what is useful amid the search for a larger coherence in our sciences, in our understanding.
We evolved in a context of multiple forms of cooperation (Toamasello, 1999), but other motives also anchor our expression, deception being one. But we are by nature a cooperative species: discerning relationships is key to our cephalic capabilities.
Foraging for coherence is dependent upon others. We discern reciprocal relationships with the investigators and are guided by them into diverse worlds in which we practice, perform, and survive. The ambiance of neuroscience – its very practice – is forever linked to dependence on others in being guided by, and by guiding others into, action in a sea of joint collaborative bonds. We come prepared for this. Even infants can direct, inform, and reciprocate (Kagan, 1984). Stick your tongue out at even a day-old baby, and the child will respond after a few times with the same gesture; the infant will later initiate the game. The collaborative bond is fundamental in order for the child to get a foothold in the world.
Tracking events and statistics
Statistics was once seen in quite pejorative terms (Hacking, 1964). An age that was misguided with regard to the notion of necessity for most biological and physical events saw statistical reasoning as a faulty form of knowledge. Isaac Newton, for instance, disparaged uncertainty in physics and relied on necessity for the prediction of the heavens. Immanuel Kant, also, made necessity a condition of what we impose by our categories for understanding.
Much of real inquiry, however, involves struggling with the unknown, the barely-known, or the yet-to-be-known (Dewey, 1925). Inquiry in neuroscience occurs between pockets of knowledge and only somewhat reliable methods of exploration. Hypothesis testing, inference, and feedback are at the heart of this process of discovery.
Indeed, from our routine problem solving for survival emerged our ability to discern and act on that which is knowable only with degrees of confidence. That judgmental practice is the heart of statistical inference. We come prepared for statistical reasoning by way of our central nervous system.
We now know that diverse regions of the brain are linked to statistical inference, and they include both neocortical and subcortical regions such as the basal ganglia (Knowlton, Mangels and Squire, 1996). Perhaps it is not surprising that cortical regions are linked to statistical inferences, but the subcortical sites may be surprising as these sites are also understood in the context of motor regions. ...
Table of contents
- Cover
- Title
- Introduction Beginnings: Foraging and Neuroscience
- 1 Foraging, Learning, and Knowing
- 2 Keeping Track of Objects in a Visual Species
- 3 Active Brains in Search and Satisfaction
- 4 Coping and Sampling in Biological Machinery
- 5 A Field Expanding
- 6 Motivation, Endurance in an Adaptive Brain
- 7 Some Therapeutic Expectations, Strategies, and Alternatives
- Conclusion: Neuroscience, Experience, and the Larger Culture
- References
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Pragmatism and the Search for Coherence in Neuroscience by Jay Schulkin in PDF and/or ePUB format, as well as other popular books in Philosophy & Mind & Body in Philosophy. We have over 1.5 million books available in our catalogue for you to explore.