Sauropods, those huge plant-eating dinosaurs, possessed bodies that seem to defy every natural law. What were these creatures like as living animals and how could they reach such uniquely gigantic sizes? A dedicated group of researchers in Germany in disciplines ranging from engineering and materials science to animal nutrition and paleontology went in search of the answers to these questions. Biology of the Sauropod Dinosaurs reports on the latest results from this seemingly disparate group of research fields and integrates them into a coherent theory regarding sauropod gigantism. Covering nutrition, physiology, growth, and skeletal structure and body plans, this volume presents the most up-to-date knowledge about the biology of these enormous dinosaurs.

- 344 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
PART ONE
NUTRITION
2
Sauropod Feeding and Digestive Physiology
Sauropod dinosaurs dominated the large herbivore niche in many Mesozoic ecosystems. On the basis of evidence from extant herbivores, significant symbiotic gut microbe activity can safely be inferred for these animals. A hindgut fermentation chamber as in horses or elephants appears more likely than a foregut system. Sauropods are unusual in several herbivore-relevant features such as their large foraging range (due to a long neck), apparent lack of food comminution (which is highly untypical for large extant herbivores), and their extremely high body weights (which is likely linked to several key features of herbivore foraging and digestion). On the basis of regressions on extant herbivores, their gut capacity can be safely assumed to have been highly comprehensive in relation to energy requirements. This can, but need not necessarily, imply extremely long food retention times. Besides these animal features, the spectrum of food plants available for sauropods in sufficient quantity (sphenophytes, pteridophytes, and gymnosperms) was completely different from that of extant herbivores (mostly angiosperms), which has some potential implications for the respective harvesters of these plants. Gymnosperms have a tendency to facilitate rather large cropping sizes (measured in kilograms of dry matter per bite) and therefore large intakes. In vitro digestibility of several living representatives of potential sauropod food plants was estimated to be better than expected, and at least comparable to the level of extant browse. Although sauropods are different from extant large herbivores in several aspects, they must be considered one of the greatest success stories in the long history of large animal herbivory.
Introduction
The longnecks, as sauropods are sometimes called by young dinosaur enthusiasts, are still often perceived as gigantic but strange creatures with a funny body shape, rather than as evolutionary successful animals. However, because they are the largest herbivores ever, as well as the terrestrial vertebrates that dominated the megaherbivore niche of most land masses from the end of the Triassic until the end of the Cretaceous for an incredible 135 million years, they should instead be regarded as the most successful vertebrate herbivores ever known. When referring to them as ‘‘the sauropods,’’ one must not forget that this group is made up of a large group of diverse herbivores that should probably be no more regarded as uniform in their digestive physiology than, for example, ‘‘the primates,’’ which also utilize a great variety of digestive strategies.
The differences repeatedly demonstrated in the skull anatomy and dentition between different sauropod clades (Calvo 1994b; Christiansen 2000; Upchurch & Barrett 2000), for example, exceed in their complexity those observed between artiodactyls and perissodactyls. It seems likely that some taxa were specialized at least to some degree on certain groups of plants—just like many megaherbivores today that can feed quite selectively on certain plant types such as grass (hippo, white rhino, large bovids), browse (giraffe, black rhino, Sumatran rhino), or use a combination of grass and browse (Indian rhino, with some tendency to include more grass in its diet, or elephants, of which African elephants have the tendency to include less grass in their diet compared to the Asian elephant) (Clauss et al. 2008a). When speculating on the digestive physiology of sauropods, one has to be aware that we rely almost exclusively on extrapolations from extant organisms; in other words, we have to extrapolate far beyond the body mass range from which our knowledge on digestive processes is derived, and we have to use as modern analogs another clade (mammals) that currently occupies the megaherbivore niche. Almost all information on the digestive physiology is in the soft tissue of the stomach and intestines, which do not occur in the fossil record of dinosaurs. Although a variety of coprolites and fossilized gut contents have been described from herbivorous dinosaurs (Stokes 1964; Chin & Gill 1996; Hollocher et al. 2001; Ghosh et al. 2003; Prasad et al. 2005), hardly any of these can be safely considered of sauropod origin (Sander et al. 2010); therefore, this source of information is at the moment not available for sauropod research. Instead, we have to speculate on the digestion of sauropods by means of educated guesses that are in part based on extrapolations from extant herbivores.
Just about everything has been said about dinosaur feeding (Coombs 1975; Bakker 1978; Krassilov 1981; Weaver 1983; Coe et al. 1987; Farlow 1987; Dunham et al. 1989; Weishampel & Norman 1989; Dodson 1990; Taggart & Cross 1997; Tiffney 1997; Upchurch & Barrett 2000; Magnol 2003). Any attempt on our part to outline the physiological characteristics of dinosaurs will therefore by necessity reiterate statements that can be found somewhere in the scientific literature. Especially because we are dealing with speculation rather than hard data, it is often difficult to properly honor all those who have already published a similar thought. For this reason, our review is not meant to be a conclusive history of all citations and ideas but instead a selective presentation. However, we want to specifically mention here the insightful works by Farlow (1987) and Paul (1998) that touch on sauropod feeding, nutrition, and digestive physiology.
Sauropods are different. Apart from our fascination in reconstructing these giants, they show alternative evolutionary strategies in vertebrate digestive physiology that we would have not thought of, and by doing so, they elucidate constraints under which extant herbivores operate that we would not have noticed as constraints but rather would have taken as a matter of course.
Feeding and Food Processing in Sauropods
On the basis of analyses on their dentition, all sauropods appear to have been exclusively herbivorous as adults (Upchurch & Barrett 2000; Weishampel & Jianu 2000; Barrett & Upchurch 2005; Stevens & Parrish 2005; Sander et al. 2009, 2010a), which does not exclude the occasional use of arthropods or other small animals by hatchlings (Barrett 2000).
NECKS
Among the most remarkable features of sauropods are their large body size and very long necks. Neck types and body forms vary in sauropods, with different types such as Brachiosaurus (long neck with long front legs), Diplodocus (long neck but rather short front legs), or Dicraeosaurus (rather short neck). The biomechanical function of the neck (Stevens & Parrish 1999, 2005; Christian 2002; Christian & Dzemski 2007; Christian & Dzemski, this volume) and evolutionary causes behind the evolution of a long neck (such as that hypothesized for giraffe by Simmons & Scheepers 1996; e.g., sexual selection, Senter 2007) have received considerable attention and discussion (for a review see Sander et al. 2010b). Despite its apparent obviousness, it has only recently been shown explicitly for the giraffe that its long neck is most likely an outcome of feeding competition within the browsing guild (Cameron & Du Toit 2007). Du Toit (1990) also described a clear stratification of feeding height between giraffe and other African browsing ruminants. It appears most likely that this interpretation applies to sauropods as well—a result that sorts sauropod taxa according to their feeding height (Upchurch & Barrett 2000). Given their enormous neck, the feeding range of most sauropods has to be regarded as extremely large, a characteristic also found in the elephant, the largest living herbivore, as a result of its trunk (Colbert 1993).
SKULL AND TEETH
The pencil-shaped teeth restricted to the front of the snout in diplodocoids and titanosaurs versus the more massive dentitions of spoon-shaped teeth with wear facets in basal sauropods and basal macronarians can be regarded as extremes of sauropod dentition and skull types (Fig. 2.1). Although the former type suggests a raking type of plant cropping (the animal raking off the leaves of a twig, leaving behind the stripped, less digestible woody shoots), the latter type of teeth allows some biting off and potentially a limited degree of mastication in the sense of puncturing or even crushing the material—or at least damaging the leaf cuticle, the major barrier for microbial access—during ingestion. Differences in the microwear of teeth have been demonstrated (Fiorillo 1998), and corresponding differences in selected food type have been proposed (Bakker 1986; Galton 1986). Recent investigations and finds reveal an unexpected diversity of dentitions (Barrett & Upchurch 2005), and a more detailed separation of skull types can be applied that produced varying degrees of oral processing of food (Calvo 1994b; Christiansen 2000; Upchurch & Barrett 2000; Barrett & Upchurch 2005). In our opinion, the term ‘‘oral processing’’ should be avoided because it is ambiguous; it is not clear to what extent biting off/cropping of forage, or masticating/comminution of the cropped forage material is meant. More descriptive terms, such as ‘‘biting off, ‘‘ ‘‘stripping off, ‘‘ ‘‘chewing,’’ ‘‘mastication,’’ and ‘‘particle size reduction,’’ and even the more self-evident components of oral processing such as swallowing and lubrication, would facilitate a better understanding. Whatever component of oral processing is referred to in the literature, compared to mammals, sauropods are exceptional herbivores insofar as their teeth lack any adaptation for masticating and grinding food. In concert with different feeding heights, differences in dentition and the extrapolated way of cropping food are commonly thought to have contributed significantly to niche separation of sympatric sauropod taxa, although a concrete interpretation of what the different feeding niches might have consisted of remains vague.
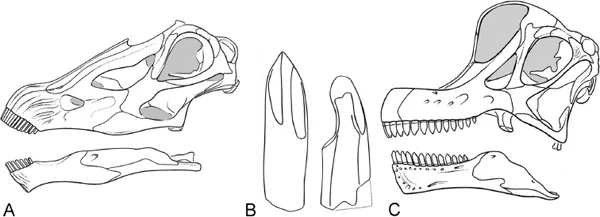
FIGURE 2.1. Sketch of two different sauropod skulls and their characteristic dentition. (A) Skull of Diplodocus sp. Note the typical small pencil-like teeth restricted to the anterior part of the skull. This kind of dentition allows only cropping and raking off plant material and no further mastication. (B) The spoon-shaped teeth of Brachiosaurus sp. are larger and show wear facets with a typical abrasion pattern.(C) Skull of Brachiosaurus sp. Note the far posterior reaching dentition. Modified after Wilson & Sereno (1998).
GASTRIC MILL
The alternative efficient option of food processing realized in extant vertebrates is the gastric mill of birds. Although gizzards with some grinding function have been described in a variety of invertebrates (Morton 1979; Dall & Moriarty 1983), among vertebrates, only some herbivorous fish such as mullets, have been reported, apart from the birds, to have a functional food particle size reduction device within their guts (Guillaume et al. 1999). Many authors have favored ...
Table of contents
- Cover
- Title
- Copyright
- Contents
- List of Contributors
- Preface
- List of Institutional Abbreviations
- Introduction
- Part 1 · Nutrition
- Part 2 · Physiology
- Part 3 · Construction
- Part 4 · Growth
- Part 5 · Epilogue
- Appendix: Compilation of Published Body Mass Data for a Variety of Basal Sauropodomorphs and Sauropods
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Biology of the Sauropod Dinosaurs by Nichole Klein in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Evolution. We have over 1.5 million books available in our catalogue for you to explore.