This volume of the series Handbook of Zoology deals with the anatomy of the gastrointestinal digestive tract – stomach, small intestine, caecum and colon – in all eutherian orders and suborders. It presents compilations of anatomical studies, as well as an extensive list of references, which makes widely dispersed literature accessible.
Introductory sections to orders and suborders give notice to biology, taxonomy, biogeography and food of the respective taxon.
It is a characteristic of this book that different sections of the post-oesophageal tract are discussed separately from each other. Informations on form and function of organs of digestion in eutherians are discussed under comparative-anatomical aspects. The variability and diversity of anatomical structures represents the basis of functional differentiations.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
21 to 23 General remarks on Erinaceomorpha, Soricomorpha and Pholidota
The Laurasiatheria (Murphy et al., 2001a, b; Kriegs et al., 2006; Hu et al., 2012) consist of the orders Carnivora, Perissodactyla, Artiodactyla and Chiroptera, which will be presented later, as well as of the following orders with genera and species, as listed by Wilson and Reeder (2005):
Erinaceomorpha (hedgehogs) with 10 genera and 24 species.
Soricomorpha (shrews) with 45 genera and 428 species.
Pholidota (pangolins) have only one genus, including eight species.
Due to the admirable efforts of Schultz (1965), there is remarkable amount of information referring to the digestive tracts of the Chiroptera, but for the other three orders, especially for the Erinaceomorpha and the Pholidota, there is no reliable comparative information with detailed illustrations, especially not for the large intestine. Information on the morphology of the digestive tract in these orders is very unbalanced. It represents an additional complication that a caecum in Laurasiatheria can be totally absent in some orders (Erinaceomorpha, Soricomorpha and Pholidota) (Tab. 5.1), or present in some and absent in other species of an order (Chiroptera, Cetacea: Odontoceti).
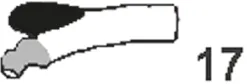
Tab. 5.1: Schematic compilation of caecal anatomy in Laurasiatheria. The numbers to the right of each drawing refer to the following literature sources: 4. Huntington (1903), 8. Jacobshagen (1937), 12. Takahashi and Yamasaki (1972), 16. Starck (1982), 17. Amasaki et al. (1989).
Order
Suborder
Caecum
Erinaceomorpha
Erinaceidae
Soricomorpha
Chiroptera
Pholidota
Manidae
Carnivora
Feliformia
Carnivora
Caniformia
Perissodactyla
Ariodactyla
Catacea
Mysticeti
Cetacea
Odontoceti
21Erinaceomorpha
21.1Introductory remarks and notes on food of Erinaceomorpha
This order consists of 10 recent genera and includes 24 species, 16 belonging to the subfamily Erinaceinae (Hutterer, 2005a), which are spiny hedgehogs, and eight to the Galericinae or moonrats and Gymnures (Gould, 1995; He et al., 2012) without spines. The latter authors consider Erinaceidae as “the oldest known living placental mammals”, “extending back to the early Paleocene of North America” (He et al., 2012, page 1). The monophyletic Erinaceinae can be found in Eurasia and Africa, the Galericinae, which are also monophyletic (Gould, 1995), live in South and Southeast Asia, as well as on Philippines and Indonesian islands. Maps showing the geographical distribution of spiny hedgehogs have been published by Reeve (1994). A short account of the history of African Erinaceidae and their two subfamilies Erinaceinae and Galericinae was published by Butler (1985).
Food of Erinaceus europaeus, the western hedgehog (Erinaceomorpha) consists largely of invertebrates at ground level, e.g. earthworms, carabid beetles, caterpillars, spiders, slugs. Occasionally vertebrates are eaten, e.g. frogs, lizards, young rodents, and nestlings as well as bird’s eggs and carrion including fish and some plants, fruits and fungi (Macdonald and Barrett, 1993). Southern African hedgehogs (Atelerix frontalis) are omnivores (Skinner and Chimimba, 2005). Invertebrates (e.g. beetles, earwigs, grasshoppers, termites, slugs, snails, centipedes, millipedes, moths and earthworms) form the bulk of their diet. They also consume small vertebrates (e.g. mice, lizards, frogs, the eggs and chicks of ground-living birds) and some vegetable matter, including fungi.
Describing the biology of Atelerix albiventris (four-toed hedgehog of East Africa, Kingdon, 1974a writes Erinaceus albiventris) that the author identifies insects, earthworms, snails and slugs as favourite food, but a wide range of other animal and even vegetable materials are also eaten, such as eggs and ground-nesting birds, small mammals, frogs, reptiles, crabs, fruit, fungi, roots and groundnuts have been reported. These findings have more recently been corroborated by the investigations of Santana et al. (2010). This means that this species has to be classified as omnivorous. Very similar data for the food of Atelerix frontalis, the Southern African hedgehog, are given by Skinner and Chimimba (2005). Erinaceus concolor, the Eastern European hedgehog, is also called “omnivore” by Qumsiyeh (1996) – they “eat almost everything given to them” (page 63); KryŠtufek and Vohralík (2001) speak of omnivory. Grosshans (1983) mentions for Erinaceus europaeus, the West European hedgehog, that beetles, Coleoptera, represent the staple food in Schleswig-Holstein (northern Germany). Plant material is only eaten accidentally; sometimes seeds and fruits are taken, but during the rest of the year they do not contribute significantly to the food. Only in autumn fruits play a role as food constituents (Holz and Niethammer, 1990a). Investigating western European hedgehogs in England Dickman (1988) found that young animals take the food from the entire spectrum of available prey types, but older animals sample prey from a narrow range of material. According to Holz and Niethammer (1990b) coleopterans are important food material in Erinaceus concolor, the southern white-breasted hedgehog. Atelerix algirus, the North African hedgehog (according to Saint Girons [1969] the latter species can also be found in southern France) prefers insects, but small reptiles can also be ingested (Holz and Niethammer, 1990c). In contrast to their appearance in the diet, adult beetles were less important in E. europaeus in total energy terms, but retained an important position along with caterpillars and earthworms (Reeve, 1994). In the subfamily Galericinae, the food seems to be similar to that of the spiny hedgehogs. For example, Sheng et al. (1999) mention that Neotetracus sinensis (authors write Hylomys sinensis) feeds “on various insects and a variety of roots and stems” (page 4).
Recently, Pereira et al. (2016) published a comparison of the gastrointestinal tracts of the rodent Jaculus jaculus, the lesser Egyptian yerboa, and Paraechinus aethiopicus, the desert hedgehog, an erinaceomoph species. The information presented in that paper is rather limited. The hedgehog eats mainly insects with some additional plant material and the surface enlargement factor in its stomach is significantly lower than in the yerboa, but for the gut (small plus large intestines), such differences are not clearly given. A caecum is absent in the hedgehog, but the other sections of the gastrointestinal tract do not show great differences between both unrelated species.
21.2Anatomy of the stomach of the Erinaceomorpha, including arterial supply
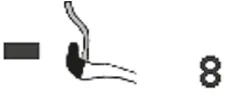
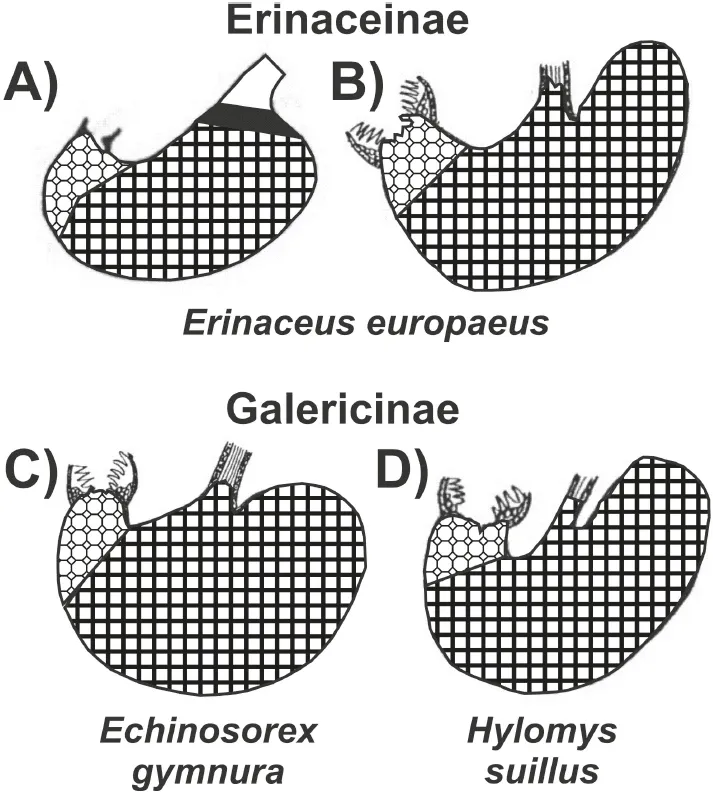
Some information is available for the gastric anatomy of Erinaceinae. According to Oppel (1896), there is no prominent incisura angularis on the lesser gastric curvature of Erinaceus europaeus. A cardiac gland zone is present, but very small and not well differentiated. Around the cardia, there is a 2- to 3-mm narrow zone of cardiac mucosa. This has been illustrated by Pernkopf (1937) (Fig. 5.1 A, black), but cannot be found in the picture supplied by Zhukova (2001) (Fig. 5.1 B). The zone of proper gastric glands (Pernkopf, 1937) extends over 90% of the total internal gastric surface; the pyloric gland zone, close to the effluent aperture of the stomach, is small.
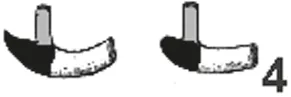
A hedgehog will eat a third of its body weight in one night (Kingdon, 1974a), according to Rossolimo (1955), fide Zhukova (2001), the daily food consumption in Erinaceus europaeus equals approximately 42% of the body weight. Erinaceidae have a voluminous and saccular stomach (Pernkopf and Lehner, 1937, Fig. 5.2) and it is possible “to consume large amounts of food in short periods of time” (Zhukova, 2001, page 393). Girgiri et al. (2015) published an illustration of the stomach of Atelerix albiventris, the four-toed hedgehog, and gave some measurements on 12 animals: The lesser curvature is considerably shorter (mean: 3.32 ± 0.37 cm) and the greater curvature is 7.42 ± 0.95 cm long, so that the organ forms a pouch-like bag. Zhukova (2001) also published illustrations of two species of the Galericinae, namely Echinosorex gymnura (moonrat, Fig. 5.1 C) and Hylomys suillus (short-tailed gymnure, D). In all four panels of the compiled illustration, the border between the proper gastric mucosa (dark rectangles) and the pyloric glands (lighter octagons) has been drawn at the aboral end of the transitional zone that was identified by Pernkopf (1937) and Zhukova (2001). Only in the drawing published by Pernkopf (1937) cardiac glands are depicted as a narrow “ring”, marked black, around the oesophageal opening.
Fig. 5.1: Gastric outlines and mucosal linings from species belonging to two subfamilies of Erinaceomorpha. The different signatures refer to cardiac mucosa (black, only in A), proper gastric and pyloric gland mucosa. Modified from: Pernkopf (1937) (A) and Zhukova (2001) (B, C, D).
Fig. 5.2: Internal aspect, showing the tunica muscularis, of the opened stomach of Erinaceus europeaus. Adapted from Pernkopf and Lehner (1937).
Cornelius et al. (1975), working on Carnivora, write that most vertebrate species that eat organisms containing chitin (e.g. insects, fungi), synthesise chitinolytic enzymes in their digestive systems, principally secreted by the gastric mucosa, in some species also by the pancreas. Chitinolytic enzymes are not of bacterial origin (Jeuniaux, 1961). In the higher vertebrates, the capacity of a given species to synthesise chitinases seems also to be related to the nature of the diet: Species which are strongly adapted to a diet entirely devoid of chitin, do not secrete chitinases in their digestive system (Cornelius et al., 1975). On the other hand, species with chitinases in the gastric mucosa, such as the western hedgehog in Europe, can potentially use chitin as a source of dietary “fibre” (Jeuniaux, 1962b). Graffam et al. (1998) determined in the four-toed hedgehog, Atelerix albiventris, that the diet dry matter of the food obtained 25% as fibrous material from chitin. A food with a high load of “fibrous” component is highly indigestible, but can be degraded because of the chitinase in the digestive tract, especially in the stomach. In addition, the stomach provided the only site for digesta retention (Clemens, 1980), and thus might enable chitin degradation through gastric chitinases.
21.3Small intestine and colon in Erinaceomorpha
The morphology of small intestine and colon in hedgehogs is not sufficiently documented. It is difficult, and in most cases even impossible, to differentiate these sections of the gut that can be so easily identified in humans and many other mammals. Recently, Girgiri et al. (2015) published a short and cursory study intended to provide “a baseline on the morphometry of the visceral organs” (page 32). The small and large intestines seem to differ in diameter according to their names – the diameter of the small intestine is slightly less than that of the colon. As has already been mentioned in the introductory words on Erinaceomorpha, the border between both intestinal sections is not marked by the presence of a caecum. This fact has already been indicated by an older, but now invalid taxonomy, where Lipotyphla (including hedgehogs) were characterised by the absence of a caecum. In addition, information on the colon of Erinaceomorpha is from the publication of Mitchell (1905). The hindgut consists of a short, straight, and rather wide rectal portion. Zhukova (2001) believes that the ancestral state of the intestinal differentiation in Erinaceomorpha is indicated by the presence of villi in the large (!) intestine.
21.4Blood vessels supplying or draining the colon of Erinaceomorpha
Information on the vascular supply or drainage in the Erinaceomorpha is...
Table of contents
Cover
Title Page
Copyright
Dedication
Contents of volume II
V Laurasiatheria
VI General discussion
Literature
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Comparative Anatomy of the Gastrointestinal Tract in Eutheria II by Peter Langer, Andreas Schmidt-Rhaesa in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Entomology. We have over 1.5 million books available in our catalogue for you to explore.