![]()
CHAPTER 1
ARCHITECTURE OF THE SOIL WORLD
We live on the rooftops of a hidden world
Peter Farb 1959
It is difficult to visualize the world of the soil as it appears to a worm or a woodlouse, a mole or a microbe. We may dig a pit in a woodland, grassland or arable soil and describe the different sections exposed to view; or feel the distinctive textures of a peaty moorland soil and a sandy heathland soil. We can measure the sand, silt and clay contents, or analyse a soil for its available plant nutrients – nitrogen, phosphorus and potassium. From our perspective, such apparent abstractions are a necessary step towards understanding a soil, but this is a long way from knowing how an individual plant rootlet will behave as it grows; or how a parasitic eelworm makes its way through the soil to attack the growing root.
Partly, it is the three dimensionality of the soil environment, and partly its physical complexity and scale, which are beyond our direct experience. Quite apart from gravel and larger pieces of rock, there is more than a thousand-fold range in size between the two extremes of the spectrum of what soil scientists call ‘fine earth’: between coarse sand particles, up to 2mm in size, and those of clay minerals which are less than 0.002mm. (The sizes of the soil inhabitants cover an even larger spread, between a 200mm earthworm and a 0.002mm microbe, for example). These mineral particles, together with an intimate mixture of living and dead plant material, form a spongy matrix permeated by pores filled with air and water. The pores themselves may comprise 30–50% of the total volume in a good topsoil – plenty of space for airbreathing creatures of all sizes, and for those dependent upon an essentially aquatic way of life.
THE SOIL SURFACE AND SUPERFICIAL PLANT REMAINS
The soil environment can be looked at from many viewpoints: as a sequence of approximately horizontal zones of distinctive character and properties from the soil surface downwards; as a medium that provides varying levels of physical and nutritional support to plants with differing requirements; as a fabric affording many and varied niches suitable for particular soil organisms; as a reactive skin covering much of the earth’s surface that provides a sink and a buffer for rainfall and for airborne chemicals; and, overall, as a vital resource that sustains life on earth.
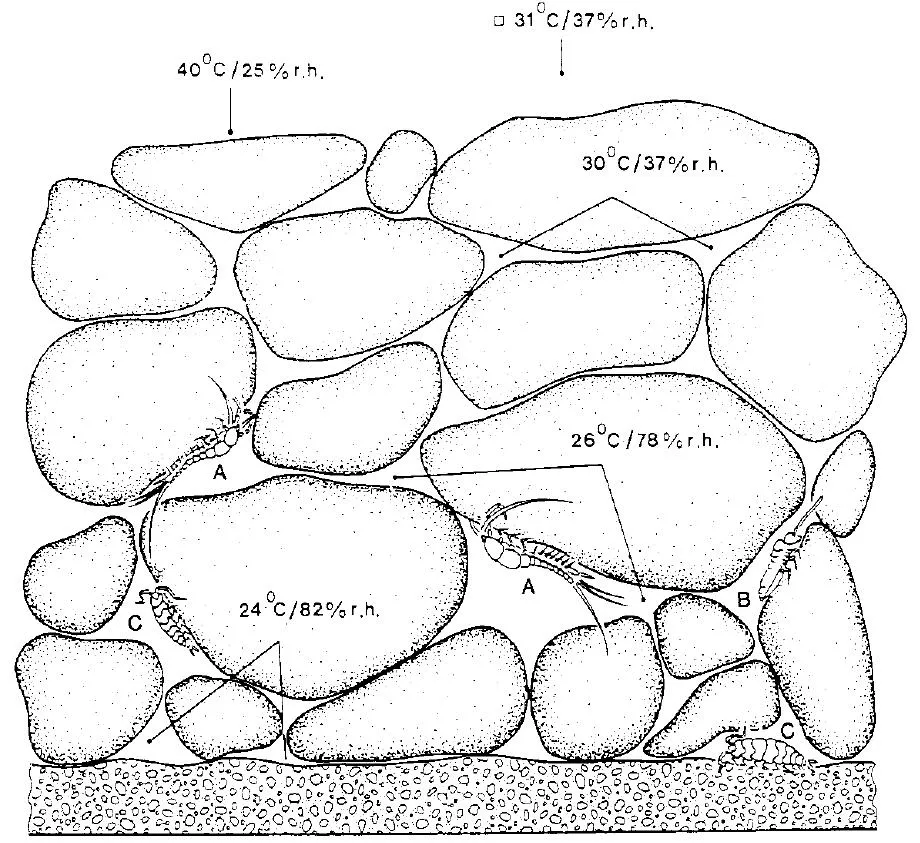
These approaches are returned to in future chapters, but in considering the architecture of the soil world it is convenient to start on its roof – at the soil surface – for this is the part that is easiest to observe, and hence is most familiar. A variety of creatures are found simply by turning over stones and logs, and while some of these may not strictly be called soil animals, yet they are often very dependent upon the nature of the underlying material at some stage of their life. Many are nocturnal, and merely shelter here during the day to avoid desiccation or predation by birds. These include predatory ground beetles and spiders, and vegetarians such as woodlice and slugs. Stones that are smaller than about 10cm are of little value for providing special microclimates unless they are scattered quite thickly over the ground as in shingle, in a quarry or on a mountainside; Figure 1 shows the temperature and moisture regimes at different depths within a heap of stones inhabited by various arthropods. Large boulders, on the other hand, are usually well bedded into the ground, and if turned over will reveal true subterranean animals such as worms, or the underground galleries of ants.
FIG. 1
Microclimate: temperature and humidity gradients in a stony habitat in strong sunlight, and the effects on positions occupied by various small arthropods; A = bristle-tail, B = springtail, C = woodlouse. The temperature decreases from the top downwards while the relative humidity increases. When the stones are shaded, the bristle-tails move to the underside of the upper stone layer. (Adapted from G. Eisenbeis 1983.)
Cracks and fissures in the ground serve much the same function as stones in affording protection from dry conditions for species that cannot burrow for themselves. Cracks are a common feature of clay soils during the summer, and may penetrate a foot or more in grassland or arable fields.
Plant cover affects conditions on the soil surface in more ways than inorganic objects do because of its more varied and complex structures and thermal properties. Adhering vegetation, such as moss and algae, liverworts and lichens, harbours a rich micro-fauna of protozoa, nematodes, tardigrades, small mites and springtails. Larger moss cushions, grass tufts and rosette plants shelter a wealth of small beetles and other insects, both adults and larvae. Dead plant material not only gives shelter but offers food for a wide range of animals and fungi which fulfil a vital role in returning the store of organic material and nutrients back to the soil.
FIG. 2
Leaf litter, twigs and branches in a mixed oak/ash woodland in spring. Note that oak leaves remain but all the ash leaves have disappeared. (Photograph B.N.K.D.)
A forest floor has the greatest variety of plant litter – annual sheddings from the tree canopy with occasional branches and logs, all in varying stages of disintegration and decomposition (Fig. 2). The total litter fall in a deciduous woodland in this country is around 2.5 kg a square metre (10 tons an acre). Seventy-five percent of this is leaf litter, the rest being made up of twigs, bark and seeds etc. The detailed character of this litter is important in providing habitats and food sources for different organisms. In a particular study of woodland litter, H. Heatwole decided there were three main categories. His first class consisted of leaves that roll or bend when they fall to the ground, thus producing large, round or angular spaces between them. Class 2 consisted of leaves that remain flat and so have small, narrow interspaces, and class 3 consisted of solid, woody objects. Each class was subdivided, for example to distinguish thick leathery leaves from thin papery ones and conifer needles, or accumulations of twigs from large logs. Some of these types are characteristic of natural, mixed deciduous woodland, beech hangers or pine woods in this country. A few are perhaps associated more with parks and gardens, where exotic trees and shrubs like rhododendron have been planted.
There is an enormous difference in the persistence and smothering effect of different kinds of leaves, ranging from ash and apple, which disappear in weeks, to beech, holly, rhododendron and conifer needles which may last for years. This difference is partly due to their size and thickness, and partly to their palatability to soil animals and susceptibility to fungal attack. The depth of litter can vary greatly depending on the density of trees, the time of year, the properties of the underlying soil, and the micro-relief of the ground. Mounds and convex surfaces may remain largely bare of litter while leaves and twigs accumulate in hollows which therefore act as foci for litter-seeking invertebrates. The student of these groups soon gets an eye for such ‘hot spots’; with experience, he can judge very accurately what species to expect.
Where litter persists for several months or years, one can usually see three distinct organic layers above the mineral soil itself. The uppermost layer of curled and uncompressed leaves has a great deal of interstitial space. This is the zone favoured by large, active springtails which grow to 5-6mm in size, and which form the prey of many beetles and spiders. Both hunting and web-building spiders exploit this open-textured but sheltered environment. The webs may be fairly simple arrangements of criss-cross threads spun across the ends of the rolled leaves, but these suffice to entangle or delay weak prey. Experiments have shown that leaf characteristics influence the numbers of spiders in woodland litter: curled leaf litter tends to support higher densities and a greater assortment of spiders.
Beneath this layer of relatively unaltered plant remains comes a zone of partly decomposed but still clearly recognizable plant fragments, and below this a zone of amorphous, finely divided organic matter. These three layers have been given a variety of names, but it is convenient to refer to them as the L (litter), F (fermentation) and H (humus) layers. The fermentation layer is where most of the litter decomposition takes place. This is the home of several kinds of millipedes, woodlice and fly larvae, some small earthworms, many mites and shorter-springed springtails. These are described in more detail in chapters 4 and 5.
Grassland litter differs from woodland litter in that the dead grass does not fall to the ground in the same way but remains arched over the surface for some months, and only gradually sinks down and disintegrates. Highly siliceous grasses, such as tor grass Brachypodium pinnatum, form a distinctive and persistent litter mat which few invertebrates appear able to digest. Even palatable meadow grasses, however, can give rise to a peaty mat on the surface of the ground if earthworms are absent; this was seen in New Zealand, for example, when settlers first converted the native vegetation into pasture land with introduced grasses from Europe.
We can move one step nearer to appreciating the structural diversity of litter if we take vertical sections and view them through a microscope. This has been done in both woodland and grassland by cutting small cores or blocks, impregnating them with gelatine, and slicing them up (Fig. 3 and Plate 1). In a study of woodland soils, J.M.Anderson recognized up to seven main classes of structures in a section through the litter, fermentation and humus layers of a sweet chestnut stand. These included not only various leafy and woody items and the cavities between them, but also plant roots with or without their fungal associations, faecal pellets of various invertebrates, and animal remains. He subdivided several of these classes to produce a total of 24 microhabitat categories which were thought to be significant for soil mites (Table 1). By examining many sections in a standard way, it then became possible to relate the number of kinds of mites identified in a section with an index of diversity computed from the number of microhabitats present. The richest zone tended to be the fermentation layer which attracted about 21 species of oribatid mites, whereas the humus layer below only supported some 10 species.
FIG. 3
Section through soil humus impregnated with agar jelly showing the ‘primitive’ insect Campodea staphylinus (Diplura). This species is colourless and blind but has highly developed tactile senses. It does not burrow but moves through the soil cavities using its antennae to locate a pathway when moving forward, and its equally well-developed posterior feelers’ (cerci) when moving backward under confined conditions. (Photograph J.M. Anderson.)
This approach is clearly an advance over macroscopic analyses of litter habitats but many important criteria are still left out. We could perhaps identify the kinds of leaves present, up to a certain stage of disintegration, but their chemistry and relative palatability – the presence of sugars, cellulose, waxes, tannins and lignin – would still elude us. It is as if we tried to distinguish between caster sugar and salt by eye alone.
HUMUS
We should think of this surface litter and decomposing organic matter, not just as an inert physical habitat for mites and other organisms, but more like the house which Hansel and Gretel found. This, you will remember, was made of ginger-bread, chocolate and barley sugar. The various components of fresh litter – fruits, leaves, stems and bark – differ greatly in their chemical make-up, and these differences are reflected in their rates of breakdown. The soft parts of leaves, containing sugars, proteins and starch within the cells, are quickly attacked and digested by earthworms, millipedes, springtails and other soil animals. One can often find a perfectly skeletonized leaf in which every vein has been left intact after removal of the lamina by micro-arthropods. The veins and other more woody structures are largely composed of cellulose. Snails are among the few animals that can secrete cellulase and so digest cellulose directly. Most cannot digest cellulose until it has been chemically shredded by microbial attack into simpler molecules. A few animals, ranging from termites to cattle, have evolved the trick of employing microflora in their gut for this purpose. The toughest woody fibres are composed largely of lignin which is highly resistant even to microbial attack, and these therefore remain intact for a long time. Recalcitrant, too, are waxes and resins, as can be seen in the persistence of holly leaves and pine needles.
TABLE 1
Microhabitat categories used in soil structure analysis (Adapted from J.M.Anderson, 1978)
| 1. Intact leaves | 13. Other macrophyte material |
| 2. Leaf fragments ≥5mm | 14. Clear fungal threads |
| 3. Leaf fragments ≥1mm <5mm | 15. Brown fungal threads |
| 4. Leaf fragments <1mm | 16. Faeces: free living macrofauna |
| 5. Humus/soil organic matter | 17. Faeces: plant-inhabiting macrofauna |
| 6. Twigs | 18. Faeces: free living mesofauna |
| 7. Wood fragments ≥5mm | 19. Faeces: plant-inhabiting mesofauna |
| 8. Wood fragments ≥1mm <5mm | 20. Animal remains |
| 9. Wood fragments <1mm | 21. Mineral material |
| 10. Roots without fungi (mycorrhiza) | 22. Cavities ≥5mm |
| 11. Roots with living mycorrhiza | 23. Cavities ≥lmm <5mm |
| 12. Roots with dead mycorrhiza | 24. Cavities <1mm |
The role of soil microorganisms in plant decomposition is described in chapter 6. At a simple level, the relative rates of breakdown of plant structures reflect the ratios of carbon to nitrogen in their chemical make up. Grass leaves have a carbon : nitrogen ratio of about 5:1, barley straw about 60:1 and pine needles about 100:1. Since soil microbes have a low carbon : nitrogen ratio of 7 to 6 or even 4:1, it follows that they cannot fully exploit plant tissues that have higher proportions of carbon without drawing on other sources of nitrogen. This has practical implications in the case of straw that is incorporated into the soil after harvest, a point discussed further in chapter 8. Similar arguments apply to the carbon : phosphorus and carbon : sulphur ratios, though the supplies of phosphorus and sulphur are not so limiting.
Humus is the final product of organic matter decomposition. It is a dark amorphous material consisting of complex organic molecules which can be broken down into humic and fulvic acids. There are, therefore, three interrelated organic fractions in soil. First, there are the plant (and animal) residues which form the main source of available nitrogen, phosphorus and sulphur for new plant (and animal) growth. Secondly, there is the microbial biomass which acts as a temporary store of such nutrients, and thirdly, a persistent humus fraction which is highly resistant to further breakdown but which can release nutrients very slowly. Measurements have been made of the age of humus by radio-carbon dating techniques; that is, by measuring the proportion of the radio-active isotope 14C left in the humus, and calculating the time since it must have been taken up by the living plant as carbon dioxide from the atmosphere. Such measurements give periods of several centuries, components such as humic acid persisting for over a thousand years in some instances. To use...