The fossil record on Mesozoic mammals has expanded by orders of magnitude over the past quarter century. New specimens, some of them breathtakingly complete, have been found in nearly all parts of the globe at a rapid pace. Coupled with the application of new scientific approaches and techniques, these exciting discoveries have led to profound changes in our interpretation of early mammal history.
Mesozoic mammals have come into their own as a rich source of information for evolutionary biology. Their record of episodic, successive radiations speaks to the pace and mode of evolution. Early mammals were small, but they provide key information on the morphological transformations that led to modern mammals, including our own lineage of Placentalia. Significant and fast-evolving elements of the terrestrial biota for much of the Mesozoic, early mammals have played an increasingly important role in studies of paleoecology, faunal turnover, and historical biogeography. The record of early mammals occupies center stage for testing molecular evolutionary hypotheses on the timing and sequence of mammalian radiations.
Organized according to phylogeny, this book covers all aspects of the anatomy, paleobiology, and systematics of all early mammalian groups, in addition to the extant mammalian lineages extending back into the Mesozoic.

eBook - ePub
Mammals from the Age of Dinosaurs
Origins, Evolution, and Structure
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Mammals from the Age of Dinosaurs
Origins, Evolution, and Structure
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
Physical SciencesSubtopic
BiologyCHAPTER 1 | |
 | Introduction |
The first 155 million years (Ma) of mammalian history occurred during the Mesozoic. During this vast time span, mammals diversified into many lineages and underwent enormous anatomical evolution. Humans are primates; primates are placentals; placentals are eutherians; and stem eutherians have a long history, which extends well back into the Early Cretaceous. Mesozoic mammals include the trunk and a bewildering bush of basal branches for the entire mammalian family tree. Their fossil records are indispensable for our understanding of the deep history that gave rise to extant mammals, including our own lineage of placentals.
Mammals have thrived in the world’s biota for 65 Ma, since the beginning of the Cenozoic, and they are represented today in a spectacular diversity of more than 4,600 modern species (Nowak, 1991; Wilson and Reeder, 1993; MacDonald, 2002). These diverse Cenozoic and modern mammals are descendants of three major evolutionary lineages: eutherians (including placentals), metatherians (including marsupials), and australosphenidans (including monotremes). The surviving members of these lineages, however, represent only a tiny fraction of a dazzling array of some 25 distinctive mammalian clades (at the order or family level of the traditional Linnaean taxonomy) that existed in the Mesozoic. Modern mammals represent merely the tips of three twigs of a vast evolutionary bush, most of which was pruned by extinction in the Mesozoic (Clemens et al., 1979; McKenna and Bell, 1997; Cifelli, 2001; Luo et al., 2002).
PREMAMMALIAN SYNAPSIDS
The synapsid lineage that includes mammals split from other amniote vertebrates over 300 Ma ago, during the Late Carboniferous (Pennsylvanian) in the Paleozoic. Synapsids are characterized by the presence of a lower temporal fenestra in the skull, among other apomorphies. Their stem taxa, or pelycosaurs, thrived in the Late Pennsylvanian through the Early Permian (Kemp, 1982; Carroll, 1988; Hopson, 1994). Therapsids, a derived subgroup of synapsids, are characterized by enlargement of the temporal fenestra to accommodate enlarged jaw muscles, a distinctive notch in the angular bone of the lower jaw, several derived features in the limb girdles, and a more anteroventral (near parasagittal) posture of the hind limbs than in the pretherapsid synapsids. Therapsids first appeared at the beginning of the Late Permian and promptly split into several stem groups, which dominated the terrestrial biota in the Late Permian. Cynodonts, a derived subgroup of therapsids, appeared in the Late Permian. Cynodonts are distinguished from precynodont therapsids in having a better-developed secondary bony palate, to separate the nasal passage for breathing from the mouth cavity for feeding, and a larger coronoid process on the dentary, with a definitive masseteric fossa for strengthened jaw adductor muscles, among many other derived features (Kemp, 1982; Hopson, 1994). The advanced cynodonts, such as probainognathids, tritylodontids, and tritheledontids, achieved many mammal-like features in both the skull and postcranial skeleton (see chapter 3).Mammalia (see box, next page) are nested within the advanced cynodonts and are defined here as all descendants of the common ancestor of Sinoconodon and extant monotremes, marsupials, and placentals. Mammals so defined are diagnosed by the dentary condyle to the squamosal glenoid in the jaw hinge and the presence of the petrosal promontorium. The earliest-known and transitional taxa bearing these and other derived features appeared about 220 Ma ago, in the Norian age of the Triassic, possibly as early as the Carnian (Fraser et al., 1985; Lucas and Luo, 1993; Sigogneau-Russell and Hahn, 1994; Datta and Das, 1996; Jenkins et al., 1997; see also figure 1.1). The poorly known Adelobasileus Lucas and Hunt, 1990, is also tentatively included within the Mammalia.
A substantial body of evidence supports the hypothesis that therapsids achieved a higher level of overall activity and more elevated growth rates than pretherapsid synapsids. Brink (1956, 1980) speculated on the basis of skeletal evidence that some of the nonmammalian cynodonts (advanced “mammal-like reptiles”) acquired hair and a diaphragm, which he suggested are indicative of an evolutionary trend toward endothermy. Bakker (1971, 1975) argued that predator-prey ratios of Late Permian–Early Triassic therapsid communities were closer to those of extant endothermic communities than to ectothermic communities. Based on a study of bone histology, Ricqlès (1974, 1976) concluded that therapsids had a high level of overall activity and high growth rates in comparison to pelycosaurs. He argued that nonmammalian therapsids probably had high metabolic rates and were likely to have achieved some kind of endothermy. In comparison to the pretherapsid pelycosaurs, advanced nonmammalian therapsids had a more upright posture, indicating a more active locomotory pattern (Kemp, 1982; Hopson, 1994). Several studies (e.g., Bennett and Ruben, 1986; Hillenius, 1992, 1994) have documented the presence of a maxillary ridge in the nasal cavity in nonmammalian therapsids. This provides indirect evidence for the development of maxillary turbinates, which in turn are related to thermoregulation in extant mammals (Moore, 1981). Hillenius (1994) proposed that thermoregulatory adaptation was developed among cynodonts and even in precynodont therapsids. Although there is broad consensus that therapsids had elevated levels of overall activity and growth rates, it remains unclear as to whether any nonmammalian therapsids (including cynodonts) developed the same homeothermic (constant body temperature) physiological functions as those seen in modern mammalian insectivores (Crompton et al., 1978).
Mammalia are a clade defined by the shared common ancestor of Sinoconodon, morganucodontans, docodontans, Monotremata, Marsupialia, and Placentalia, plus any extinct taxa that are shown to be nested with this clade by parsimony analyses. This stem-based definition excludes from Mammalia such advanced cynodonts as chiniquodontids, probainognathids, tritylodontids, tritheledontids, or any taxa nested within a subset of the latter cynodont groups. The Mammalia concept here is equivalent to the Mammaliaformes of Rowe (1988: figure 4, but not that of Rowe, 1993: figure 10.2).
In their impressive book, Classification of Mammals above the Species Level, McKenna and Bell (1997) adopted a crown-based concept of Mammalia, originally proposed by Rowe (1988) and subsequently accepted by some other authors. Following the definition by Rowe (1988) and McKenna and Bell (1997), several groups regarded here to be mammals (e.g., Morganucodonta, Docodonta, Haramiyoidea) would be excluded from the Mammalia, instead being regarded as nonmammalian mammaliaforms. The status of several other groups—including the best known and most diverse of Mesozoic mammals, the Multituberculata—would be ambiguous under this definition, as their phylogenetic position relative to crown Mammalia has yet to be settled decisively (see chapter 15).
There is a massive body of literature dealing with the merits and problems associated with crown-, stem-, or character-based definitions of Mammalia (see Lucas, 1992; Rowe and Gauthier, 1992; Bryant, 1994). The definition adopted by us is one of several alternatives. We prefer this inclusive definition of mammals because (1) it is consistent with wide-spread, traditional usage (e.g., Hopson, 1994); (2) it has the virtue of being relatively stable in its membership contents of both living and fossil taxa (Luo et al., 2002); and (3) it happens to be diagnosed by characters that we view as being biologically significant (see chapter 3). The first two of these criteria, at least, address a primary purpose of taxonomic nomenclature: to promote stability and commonality of usage.
SUCCESSIVE DIVERSIFICATIONS OF MESOZOIC MAMMALS
Most Mesozoic mammal species are represented only by teeth and incomplete jaws. Inadequate as they are for a comprehensive understanding of whole organisms, teeth and jaws nonetheless provide useful information about the taxonomic richness and diverse feeding adaptations of early mammals. Their records show several major episodes of diversification (figure 1.1).
The earliest diversification took place among stem mammals during the Late Triassic to Early Jurassic. From this initial burst of mammalian evolution arose several groups: haramiyidans with multirow, multicuspate, “multituberculate-like” molars; Sinoconodon and morganucodontans with “triconodont-like” molars showing three main cusps in alignment; and kuehneotheriids with triangulated, “symmetrodont-like” molars. All these mammals possessed the derived mammalian feature of the dentary condyle to squamosal glenoid in jaw joint,1 but also retained the plesiomorphous (“reptilian”) features of an articular-quadrate jaw joint and the attachment of the postdentary bones (homologues of tympanic and middle ear bones of crown mammals) to the mandible. Although haramiyidans, Sinoconodon, morganucodontans, and kuehneotheriids have almost the same mandible design (notwithstanding variations in some individual characters), these groups developed astonishingly different molar structures for different feeding adaptations.
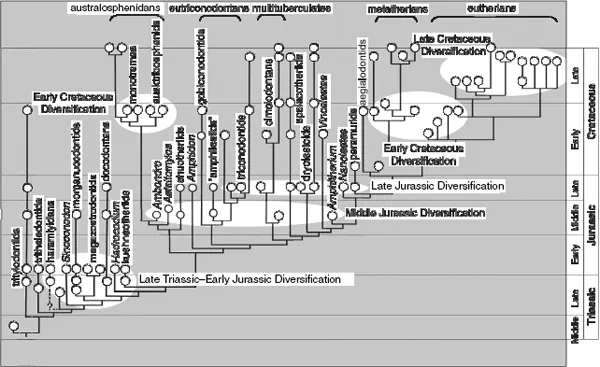
FIGURE 1.1. Overview of the temporal distribution and relationships among Mesozoic mammal clades. The great evolutionary bush of diverse Mesozoic mammalian clades is the dominating feature in their taxic evolutionary pattern. The more recent cladistic analyses of dental, cranial, and skeletal characters of all phylogenetically relevant clades have revealed a far greater array of taxa and ranks of clades than could be accommodated by the traditional views of four “long-branch” lineages to the Early Triassic (e.g., Simpson 1928a; figure 1.2A) or two prototherian versus therian divisions extending to the Rhaeto-Liassic (figure 1.2B). The almost fully resolved cladogram of all Mesozoic mammal groups, together with their improved records of temporal distribution of fossils, suggests that mammalian diversifications occurred episodically during the entire span of Mesozoic mammalian history. The five episodes of diversifications are as follows: (1) The earliest-known episode of diversification occurred in the Late Triassic–Early Jurassic on a global scale, when haramiyidans, morganucodontans, kuehneotheriids, and docodontans (if including Woutersia) appeared. (2) The next episode occurred globally in the Middle Jurassic, characterized by the appearance of shuotheriids, the earliest australosphenidans, eutriconodontans, putative multituberculates, and amphitheriid “eupantotherians.” (3) The Late Jurassic diversification occurred primarily in Laurasia among eutriconodontans, spalacotheriids, dryolestoids, and peramurans. (4) The Early Cretaceous episode saw diversification within australosphenidans and toothed monotremes on the Gondwanan continents, and the basal splits of eutherians and metatherians and diversification of tricondontids on the Laurasian continents. (5) The Late Cretaceous episode witnessed diversification within metatherians, within eutherians, and within cimolodontan multituberculates on the northern continents and of gondwanatherians on the southern continents. The earliest diversifications of stem eutherians and stem metatherians, as documented here by the currently available fossil evidence, predate the likely time window estimated by molecular evolutionary studies, which indicates that some superordinal clades of placental mammals may extend back into the Cretaceous (e.g., split of the earliest placental superordinal clades around 108 ± 6 Ma ago; Murphy, Eizirik, Johnson, et al., 2001; Murphy, Eizirik, O’Brien, et al., 2001; and several other studies). Full test of the molecular estimates of the phylogenetic divergence would require a combined analysis of currently available molecular sequence data and morphological data of major extant clades, with the main characters for resolving Mesozoic mammalian relationships coded for modern clades. This has not been done and is beyond the scope of this book. Therefore, for the time being we believe that the issue of the timing of the origin of modern placental superordinal clades is still unresolved. Source: stratigraphic distribution from chapter 2; cladogram based on Luo et al. (2002) and Ji et al. (2002).
Six more order- or family-level lineages evolved in the next episode of diversification, during the Middle Jurassic. Amphilestids, with triconodont-like molars; eleutherodontans, with multituberculate-like molars; and amphitheriids and peramurids, with a triangulate trigonid plus a talonid heel on lower molars were all present on the Laurasian continents. A major innovation of this time was that of grinding molar function, which evolved in at least three emergent lineages: docodontans, with complex shearing and grinding surfaces on the molars; shuotheriids, with an anterior pseudotalonid basin; and australosphenidans from Gondwanan continents, with a fully developed posterior talonid basin for grinding function, in addition to the plesiomorphic shearing features on the molars. It is obvious that the adaptive molar structures related to grinding functions are homoplastic among these groups.
Consistent with this is the fact that, in juxtaposition with their derived, multifunctional molars, docodontans, shuotheriids, and at least some Middle Jurassic australosphenidans retained the primitive features of the postdentary trough on the mandible. Presence of these primitive mandibular features in the Jurassic southern mammals with multifunctional tribosphenic molars is strong evidence that they are not closely related to boreosphenidans, which evolved on the northern continents some 25 Ma later, in a separate lineage with far more derived mandibular features (Luo et al., 2002).
The third episode of diversification among early mammals occurred in the Late Jurassic, with five newly emerged clades: Multituberculata (sensu stricto), triconodontids, spalacotheriids, dryolestoids (dryolestids and paurodontids), and tinodontids. The most significant apomorphic feature of these emergent clades of the Late Jurassic is the absence of the postdentary trough on the mandible (Rowe, 1993), a primitive feature retained by almost all Early and Middle Jurassic lineages (except for amphilestids and Hadrocodium). Another striking pattern concerns the structure of the dentary. Despite the fact that eutriconodontans, multituberculates, and spalacotheriids have very different dentitions, these three clades all have fairly similar characteristics in the posterior part of the mandible: a rounded “angular” region grades into the dentary condyle, and there is often a prominent medial pterygoid crest (“shelf”) along the ventral border of the mandible.
The last major episode of diversification occurred in the Early Creta...
Table of contents
- Cover
- Half title
- Title
- Copyright
- Contents
- Foreword
- Preface
- Chapter 1. Introduction
- Chapter 2. Distribution: Mesozoic Mammals in Space and Time
- Chapter 3. Origin of Mammals
- Chapter 4. The Earliest-Known Stem Mammals
- Chapter 5. Docodontans
- Chapter 6. Australosphenidans and Shuotherium
- Chapter 7. Eutriconodontans
- Chapter 8. Allotherians
- Chapter 9. “Symmetrodontans”
- Chapter 10. “Eupantotherians” (Stem Cladotherians)
- Chapter 11. “Tribotherians” (Stem Boreosphenidans)
- Chapter 12. Metatherians
- Chapter 13. Eutherians
- Chapter 14. Gondwanatherians
- Chapter 15. Interrelationships of Mesozoic Mammals
- Appendix
- References
- Additional References
- Illustration Credits
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Mammals from the Age of Dinosaurs by Zofia Kielan-Jaworowska,Richard L. Cifelli,Zhe-Xi Luo in PDF and/or ePUB format, as well as other popular books in Physical Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.