![]()
PART 1
Ligand-Gated Ion Channels
![]()
Ligand-Gated Ion Channels | 1 |
Ligand-gated ion channels mediate the rapid transfer of information across nerve synapses in the brain and peripheral nervous system. They are also located in extrasynaptic regions of neurons as well as on glia and non-neuronal tissue. These receptors are composed of four to five protein subunits that span the plasma membrane and circumscribe a central ion pore.1 When the neurotransmitter binds to its pocket on the extracellular domain of the receptor it causes the channel to open, allowing specific ions to flow in or out of the postsynaptic cell depending upon their electrochemical gradients and the selectivity of the channel. Channels that conduct an influx of sodium or calcium depolarize the cell, whereas those that conduct an influx of Cl− hyperpolarize the cell or make it more difficult to be depolarized. These channel currents instantaneously generate electrotonic currents in the region of the postsynaptic membrane. The sum of the depolarizing and hyperpolarizing currents ultimately determines the excitability of the cell. Without this rapid trans-synaptic communication, the information processing capabilities of neural networks would be severely limited.
Because ligand-gated ion channels incorporate both the ion pore and binding pocket for the chemical messenger into a single structure, it is possible to measure the activity of the ligand-receptor complex directly. This enables an unambiguous measurement of how well a drug binds to and activates the receptor. Thus, ligand-gated ion channels provide a unique opportunity to examine the properties of affinity and efficacy of the ligand-receptor complex. Any book on affinity and efficacy, therefore, must clearly explain these principles for ligand-gated ion channels. We can then apply these concepts to G protein-coupled receptors, whose activation state is not usually measured directly, but rather, inferred from measurements made at different points downstream in the receptor-signaling pathway.
This chapter focuses on the structure of ligand-gated ion channels so that we will have a better understanding of what physical properties our estimates of affinity and efficacy represent. Chapters 2–4 will describe how to measure affinity, efficacy and allosteric modulation at ligand-gated ion channels.
Ligand-gated ion channels represent a large group of receptors that can be divided into two evolutionarily related groups. One group includes receptors for acetylcholine (nicotinic acetylcholine receptors), γ-aminobutyric acid (GABAA receptors), glycine and serotonin (5-HT3 receptor).2 The other includes many types of receptors for glutamate.3 This chapter will focus on nicotinic acetylcholine and GABAA receptors. The reasons are that much has been known about the native structure of the nicotinic acetylcholine receptor for quite some time, and GABAA receptors exhibit abundant allosteric interactions. Thus, having a close look at these receptors will provide the structural basis for understanding the principles of affinity, efficacy and allosterism at ligand-gated ion channels.
Nicotinic Acetylcholine Receptors
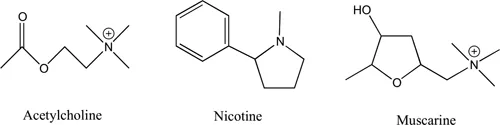
The nicotinic receptor is so-named because it represents one of the two subgroups of acetylcholine receptors that is activated by the plant alkaloid, nicotine. The other group, muscarinic receptors, is activated by muscarine, which accumulates in the fruiting bodies (basidiocarps or mushrooms) of some members of the Inocybe and Clitocybe genera of fungi. Muscarinic receptors are members of the G protein-coupled receptor family and are described in Chapter 5. Because acetylcholine is a linear flexible molecule, it can fit into the orthosteric binding pockets on both classes of receptors, whereas the more rigid alkaloids have a preference for only one type (Figure 1.1). Both nicotinic and muscarinic receptors represent groups of receptors with several subtypes.
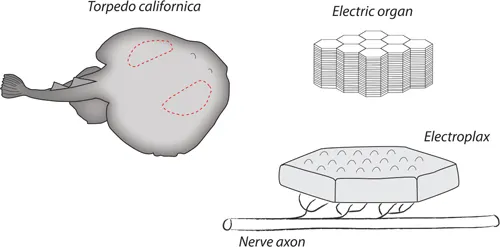
Our knowledge of the structure of the nicotinic receptor would not have progressed the way it did without a gift from nature in the form of the electric organ, particularly those from two marine rays, Torpedo californica and Torpedo marmorata.4 These fishes have large bilateral electric organs occupying a substantial portion of the flattened pectoral fins that define the skate and ray body plan. Each electric organ consists of a few hundred closely packed columns of cells traversing the ventral to dorsal sides of the ray (Figure 1.2). Each column contains a stack of about 1000 flattened cells (electroplaxs) having a diameter and thickness of approximately 5 mm by 0.02 mm. Cholinergic nerves innervate the ventral surface of each electroplax.5 When stimulated, these nerves release acetylcholine, which activates nicotinic receptors on the ventral surface of the electroplax, causing an influx of positive monovalent cations. This depolarizes the ventral side of the electroplax relative to the dorsal side causing a potential difference of about 50–100 mV. Because the 1000 or so electroplaxs in each column are linked in series and coupled in parallel with those of other columns, the total electromotive force from the discharge of the electric organ can be 50–100 V, depending upon how it is measured.
Figure 1.1:Structures of acetylcholine, nicotine and muscarine.
Figure 1.2:The electric organ from Torpedo californica. (a) The location of the bilateral electric organs in Torpedo californica is indicated by the dashed lines. (b) Schematic drawing of the columns of electroplax within the electric organ. (c) A schematic drawing of a single electroplax showing its innervation.
This large potential difference and its associated current would not be possible without the contribution of a colossal population of nicotinic acetylcholine receptors, each having a conductance of about 50 pS. This great abundance of receptors enabled Michael Raftery and colleagues6 to purify the native receptor from the electric organs of Torpedo californica, determine its pentameric subunit composition (two α1 subunits and one each of β1, γ1 and δ1), and sequence the first 54–56 amino acids of the amino terminus of the four subunits. Knowing the partial sequences, Shosaku Numa and colleagues7 determined the complete sequences by gene cloning. When injected into Xenopus oocytes, the mRNAs for the four subunits are translated into functional nicotinic acetylcholine receptors.8
Figure 1.3:Structure of single nicotinic acetylcho...