![]()
Section 1
Neuroanatomy and Physiology
![]()
Chapter 1.1
Peripheral Nervous System
Ken Horch
This chapter provides an overview of the basic structure and physiology of peripheral sensory and motor nerves.
1.Introduction
All you know about the external world, and all you can do about it, depends on your peripheral nervous system and cranial nerves, the latter a rostral extension of the former. In this chapter we provide an overview focused on the structure of peripheral nerves responsible for cutaneous and proprioceptive sensation and for voluntary muscle control.
2.Structural Organization
The peripheral nervous system (PNS) consists of two major divisions, autonomic and somatic. The autonomic system consists of a preganglionic component (with nerve cells originating in the spinal column or skull) and a postganglionic component (with nerve cells lying wholly outside the axial skeleton). The somatic system consists of nerve cells with cell bodies located within or near the spinal column or skull. The autonomic system is largely concerned with functions not normally under voluntary control, while the somatic system is concerned with sensory inputs that can be perceived and with voluntary control of skeletal muscles.
The voluntary nervous system is divided on anatomical and functional grounds into sensory and motor components. Except for the cranial nerves, the sensory system consists structurally of nerve cells with somata located outside the spinal cord in aggregates called dorsal root ganglia. These cells have no dendrites, but instead possess a single process that bifurcates upon leaving the ganglion, one process travelling centrally through dorsal roots to enter the dorsal part of the spinal cord and the other process travelling distally through peripheral nerves to innervate or form distal sense organs. These nerve processes are called afferent nerve fibers because they conduct action potentials and, hence, information from the periphery to the central nervous system.
The sensory elements can be divided, following the example of Gray [7], into the "special"senses of audition and vestibulation, smell, taste, and vision, and the "general"senses which include just about everything else that gives rise to conscious sensation. The “general” senses fall into two major domains: somatosensory and proprioceptive. One can think of the somatosensory or exteroceptive system as providing information about the state of the external environment as it interacts with the body, and of the proprioceptive system as providing information about the orientation of the body in space. This reduces to a consideration principally of cutaneous receptors and muscle/joint receptors.
The motor system consists structurally of nerve cells with somata located in the ventral quadrant (horn) of the spinal cord. These cells have dendrites ramifying through the ventral portions of the spinal cord, and single axons that leave the spinal cord through the ventral roots and join with the sensory fibers to form the peripheral nerves and eventually innervate skeletal muscle fibers. These nerve process are called efferent nerve fibers because they conduct action potentials and, hence, information from the central nervous system to the periphery.
Active movement of joints is produced by contraction of extrafusal muscle fibers. A single motor neuron will typically innervate several muscle fibers in a given muscle. Since there is normally a one-to-one relationship between an action potential on a motor neuron and an action potential in the post synaptic muscle fiber, the axon and all its innervated muscle fibers is called a motor unit. Under normal conditions, only a single nerve fiber innervates a given muscle fiber, although multiple innervation can occur transiently in response to partial denervation of muscle. The electrical properties of muscle fiber membrane makes it difficult to active these fibers by externally applied currents. Thus, except in the case of totally denervated muscle, electrical stimulation of muscle by externally applied current normally consists of activation of the terminals of motor neurons that in turn activate the muscle.
Within the spinal cord, afferent process make local synapses and may terminate within a few segments of their entry point or they may travel directly, via the dorsal columns, to the medulla of the brainstem. On the efferent side, the dendrites of motor neurons receive input from local circuits, including feedback from sensors in the skin and muscle, and input from descending pathways originating in the brain.
3.Peripheral Nerves
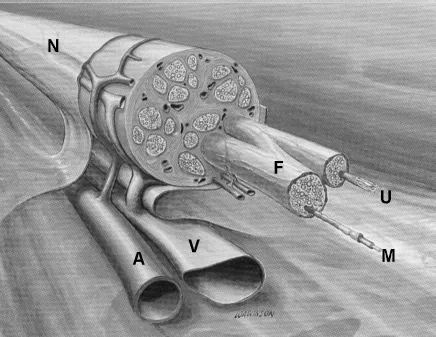
Peripheral nerves consist of the axons of autonomic, dorsal root ganglion and ventral horn neurons, their associated ensheathing Schwann cells and basement membrane embedded in a collagen matrix called the endoneurium (Fig. 1). Collections of nerve fibers are segregated by a cellular perineurium, which, along with the capillary endothelium, also acts as a blood-PNS barrier [20]. The perineurial encapsulated structure is called a fascicle. Fascicles are bound together by a collagenous epineurium, which defines the nerve as a distinct anatomical entity and is its primary provider of strength. Within their fascicles, nerve fibers follow an undulating course. This is manifest in the appearance of the Bands of Fontana, seen as alternating light and dark bands under proper illumination of the nerve. Within a given fascicle, the fibers undulate in phase. Shifts in the phase of the appearance of the light and dark bands can be used intraoperatively as an indicator of different fascicles. The undulations allow a certain amount of stretch to occur in the nerve without damage to the delicate axons. Once the nerve fibers straighten out and the Bands of Fontana disappear, further stretch of the nerve will result in damage to the axons [22].
The best sources of information about the general anatomy and mechanical properties of peripheral nerves are found in the clinical nerve repair literature, such as the classic tomes by Sunderland [22] and Mackinnon and Dellon [12] as well as more specialized review articles such as the one on mechanical properties by Topp and Boyd [23].
3.1.Nerve fibers
Nerve fibers are grouped together into fascicles not on the basis of their function, but on the basis of their destination. That is, fascicles provide a topographic organization to peripheral nerves. As one moves proximally, fascicles tend to fuse, but the topographic segregation of fibers may continue quite some distance proximally within the fused fascicle [9, 21, 27]. There is also evidence that within the somatotopic organization peripheral nerve fibers may be segregated by functional modality [4, 10]. Proximal to the cervical or lumbar plexi, the distal topographic organization is replaced by a spinal segment organization. Therefore, in order to obtain receptor specific information, one cannot rely on whole nerve or whole dorsal rootlet recording methods [2, 6, 25].
Fig. 1. Drawing of a peripheral nerve (N) with associated arterial (A) and venous (V) blood vessels. Shown as extending from the nerve epineurium are two fascicles (F) ensheathed in perineurium. Shown as extending from the endoneurium of the fascicles are a myelinated axon (M) and a bundle of unmyelinated axons encased in a single Schwann cell (U). (Adapted from Mackinnon and Dellon [12]).
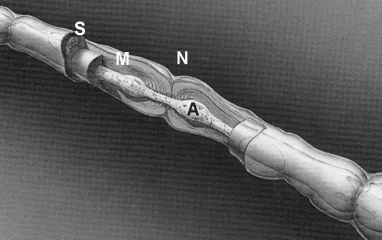
In addition to the classification of nerve fibers based on their origin and overall function (autonomic, sensory or motor), nerve fibers are also classified on the basis of the type of structure they innervate, the presence or absence of myelin, and their size. Although the conduction velocity of action potentials increases with axon size, vertebrates (unlike their invertebrate cousins such as cephalopods and annelids) utilize a second mechanism to further increase conduction velocity in large axons: myelination [28]. In these axons, the Schwann cells, rather than simply enveloping a collection of nerve fiber axons as is the case with unmyelinated fibers, form a one-to-one association with a particular axon, with a different Schwann cell every 0.5 mm or so. These Schwann cells form elaborations of their cell membranes which wrap around the axon, leaving only a small gap between Schwann cells where the nerve membrane is directly exposed to the extracellular milieu (Fig. 2). This results in saltatory conduction: action potentials jump from one node of Ranvier to the next, rather that progress continuously along the membrane as is the case in unmyelinated fibers. This results in a significant increase in conduction velocity without a concomitantly large increase in axon diameter, saving space in the nerve and metabolic requirements in the parent soma. In addition, myelinated axons tend to have briefer action potentials than unmyelinated nerve fibers, allowing them to support higher firing rates than the latter.
Fig. 2. Drawing of a myelinated nerve fiber axon (A) at the level of a node of Ranvier (N), where the myelin sheath (M) produced by the enveloping Schwann cell (S) terminates leaving a small gap before the next sheath begins. (Adapted from [Mackinnon and Dellon [12]).
In general, myelinated axons are used for tasks where speed (such as control of skeletal muscle contraction or signaling of temporally rapid and brief events) is required or where fine tactile or proprioceptive discriminations are to be made. The latter stems from the fact that the higher the signaling rate, the higher the information transfer rate. Unmyelinated fibers tend to be associated with control of smooth muscle or signaling diffuse, tempora...