![]()
Chapter 1
Perplexity of Complexity1
Systems composed of many components interacting with each other are considered complex since their behavior may not be expressed as a direct sum of individual behaviors of their parts. More complex systems naturally involve more parts and potentially integrate more diverse interactions between their components compared to simpler ones. Envisioning complex systems is an observer dependent practice [Johnson, 2014], since a knowledge of more detailed characteristics can make a system appear more complex.
Phenomena which emerge from a collection of interacting objects are often termed complex phenomena [Johnson, 2009]. The study of complex phenomena is perplexing because of emergent properties of a complex system which arise at a higher level of organization, above the level of its individual components. In this chapter we informally introduce some of the basic concepts of the modern approach to complex systems and explain why they are important.
1.1A Compositional Containment Hierarchy of Complex Systems and Processes
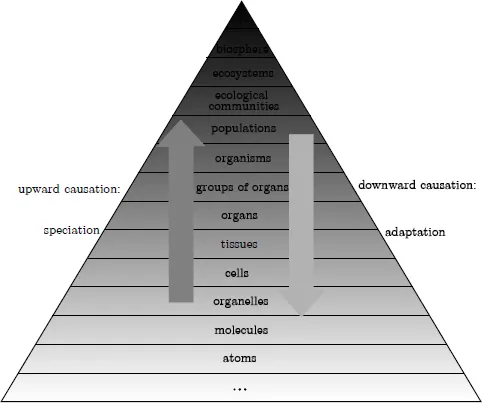
On the one hand, individual components of a complex system may be themselves complex, on the other hand the entire system embracing them may be just an elementary constituent of even more complex systems. An ordering of the parts that make up a complex system is called a compositional containment hierarchy. Each level of the compositional containment hierarchy is characterized by the certain emergent properties that may not be seen at the lower levels of hierarchy.
Atoms, the most fundamental and stable units of matter, are bonded into molecules. Aggregates of very large molecules formed by polymerization may be composed into organelles, the simplest level of organization of living things.
Different organelles may further be organized into cells. Related cells although not identical, but working together to accomplish specific functions are collectively referred to as a tissue. Tissues varying in their compositions are then joined in a higher level structural unit serving as a common function, an organ.
Groups of organs working together to perform one or more functions form organ systems. Contiguous groups of organ systems, being capable of some degree of response to stimuli, reproduction, growth, development, and homeostasis inside a defined environment, form organisms.
Collections of organisms of a species, sharing a particular characteristic and having the capability of interbreeding, are populations. Groups of populations, sharing a common environment, form ecological communities that form ecosystems functioning as a unit with their environments. The global sum of all ecosystems, largely self-regulating, forms the biosphere.
Although we stop climbing this hierarchy there, the absence of evidence is not the evidence of absence of higher levels in that. It is also obvious that the hierarchy of organization of living things is by no means a unique logical path linking the events and things in this world at the micro-, meso- and macroscopic levels.
An ordering of the parts that make up a complex system is a compositional containment hierarchy, each level of which is characterized by the certain emergent properties that are not seen at the lower levels.
On the one hand, complex systems can be fragile, since strong coupling between levels in the hierarchy can lead to cascading failures once a critical breakdown occurs in some of the system’s levels causing catastrophic consequences on the functioning of the entire system [Buldyrev et al., 2010]. On the other hand, a multi-level complex system forming a complex network of many interacting components can be extremely resilient, as being able to adapt to the rapidly changing environments [Gao et al., 2016].
1.2Top-Down and Bottom-Up Processes Associated to Complex Systems and Processes
A hierarchical organization of complex systems suggests the existence of two opposite processes linking all levels in the compositional containment hierarchy by bi-directional causation [Lane, 2006].
The compositional containment hierarchy of complex systems is characterized by bi-directional causation associated to the processes of speciation and adaptation.
1.2.1The Top-Down Process of Adaptation (Downward Causation)
Definition 1. The downward causation is a causal relationship from higher levels of a complex system to its lower-level parts [Campbell, 1974].
The corresponding top-down process enforces certain constraints from a higher level development on the behavior of lower level components of the system, making them move in ways that may be unpredictable, even the complete information about these components is given. Since the mechanisms operating at the higher levels of organization of a complex system generally fail to accomplish the tasks at the lower levels of organization directly, the top-down process of the downward causation usually takes effect indirectly, through the environment [Galaaen, 2006]. Therefore, a complex system is profoundly influenced by the environment, facing a succession of environmental challenges and actively adapting behavior to get along in the environment with greatest success.
Remark 1. Since Herbert Spencer had first used the phrase “survival of the fittest” [Spencer, 1864], we used to think that evolution is about the ability of species to adapt to the changing environment. The process of adaptation whereby a living organism is able to “leave the most copies of itself in successive generations” [Spencer, 1864] plays the central role in our contemporary understanding of the key mechanisms of natural selection favoring the species “better designed for an immediate, local environment” (as was clarified by Darwin himself in the fifth edition of his famous work On the Origin of Species published in 1869) [Freeman, 1977].
1.2.2The Bottom-Up Process of Speciation (Upward Causation)
Definition 2. The opposite (to adaptation) bottom-up process related to the upward causation involves interactions between the lower level components that causes emergent properties arising at the level of the entire system, which are not seen at the level of its basic constituents.
Emergence is particularly common when things are assembled to form new wholes [Johnson, 2014]. Emergent phenomena can occur in hierarchical systems that are far from the thermodynamic equilibrium due to a self-organization process between their parts [Prigogine et al., 1977]. The development path of a self-organizing complex system encounters bifurcation points, where the system is forced to take one of several possible ways.
The complex systems are subject to innovation, demonstrating sensitive dependence on initial conditions when a small change in the initial state of a system results in the system diverging considerably. The initially identical, but reproductively isolated biological populations can evolve to become distinct species in the process of speciation taking place over the course of evolution [Sulloway, 1982].
Remark 2. Speciation triggers the mechanism driving biodiversity and populating the levels of the pyramid shown in Fig. 1.1 with new traits, species, and ecosystems arising due to divergent natural selection among different habitats. Once identical populations become subjected to dissimilar selective pressures and/ or undergo genetic drift independently, the fixation of incompatible mutations might make them no longer capable of exchanging genes provided occasionally they come back into contact [Baker, 2005].
Fig. 1.1 A hierarchical organization of complex systems suggests the existence of two oppositely directed processes linking all levels in the hierarchy by the bi-directional causation
1.3Example: A Concept of Evolution by Natural Selection
It is the interplay between adaptation and speciation that explains the variety and variability of life on earth [Gaston, 2000].
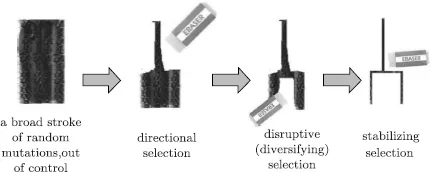
From a mathematical point of view, the theory of evolution by natural selection provides us with a recursive algorithm for drawing an infinite tree graph by a “wide brush” of random mutations and a diligent “eraser” of various selection mechanisms taking place in the normal course of adaption (Fig. 1.2).
Fig. 1.2 Evolution by natural selection represents a recursive algorithm for drawing an infinite tree graph by a “wide brush” of random mutations and a diligent “eraser” of natural selection
First, the “eraser” of directional selection is applied favoring a particular phenotype over other phenotypes independently of its dominance relative to other alleles in the past and causing the allele frequency to shift over time in the direction of that phenotype.
Second, the process of disruptive selection (also called diversifying selection) is applied favoring extreme values for a trait over intermediate values, so that the variance of the trait increases and the population appears to be divided into two distinct groups.
Third, a fine “eraser” of stabilizing selection is in use for decreasing genetic diversity and stabilizing the population mean on a particular trait value (see Fig. 1.2).
The recursive application of this algorithm allows us to draw an evolutionary tree of arbitrary size. Random mutations do not impose any particular
“stopping time” for the tree-drawing algorithm. Since most of the traits do not appear to change drastically over time, natural selection should keep on working the pitchforks out with the fine eraser continuously.
Remark 3. Despite the great success of the theory of evolution, many issues remain unexplained. In particular, we have no understanding of why some presently living organisms, such as Latimeria Menadoensis, Ginkgo Biloba, segmented worms Annelids, and many others do not undergo significant morphological changes over hundreds of millions of years, as though the factor of random mutations had no effect on them.
Furthermore, since natural selection changes the relative frequencies of alternative phenotypes and since they are heritable, it might be expected that genetic “complexity” of living species, essentially those multi-cellular, eukaryotic organisms would increase with each step of evolution. Simply speaking, the “more advanced” species are expected to be more complex as having more genes than simpler species. However, the observed reality is quite different.
For example, our Christmas tree (European spruce), an organism which is much simpler morphologically than humans, has 6.7 times as many genes as humans (2 × 1010 against 3 × 109 genes). As for the number of functional genes, the difference is even more striking, since the Christmas tree has 14.5 times as many functional genes as humans.
1.4Saltatory Temporal Evolution of Complex Systems
The dynamics of top-down and bottom-up processes in complex systems involves ...