Water is the most basic essential for plant growth; an inadequate supply causes severe problems, as plants rely on the water transmitted by soil to meet their physiological and nutritional needs. Since the first edition was published, flooding and droughts throughout the world have made water an even more topical subject, as the importance and instability of our water supplies have been brought to the forefront of daily life. This new edition of Water Dynamics in Plant Production focuses on the dynamics of water through the hydrologic cycle and the associated mechanisms that plants employ to optimize growth and development. It describes the basic scientific principles of water transport in the soil-plant atmosphere continuum, and explains the linkage between transpirational water use and dry matter production. Paying particular attention to the various agronomic strategies for adaptation to climate-driven limitations of water resources, the efficiency of water use in plant production and in achieving an economic yield is presented in detail. This book offers a multidisciplinary introduction to the fundamentals and applications of water dynamics in natural and managed ecosystems. Including text boxes throughout, as well as online supplementary material, it provides an essential state of-the-art resource for students and researchers of soil and plant science, hydrology and agronomy.This book is enhanced with supplementary resources.

- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Water Dynamics in Plant Production
About this book
Information
1 The Role of Water in Plant Life
1.1 Functions of Water in the Plant
In some ways the life of plants is much more directly dependent on water than is life in the animal kingdom. One reason for this is that higher plants differ from animals because nearly all are nutritionally self-sufficient, or autotrophic. Water serves as a hydrogen donor and thereby as a building block for carbohydrates, which are synthesized by plants making use of sunlight (Box 1.1). Another inorganic building block used by plants in the synthesis of organic primary products is carbon dioxide (CO2), which plants can only take up from the atmosphere at the same time that they return water vapour to the atmosphere. This exchange of gases is necessary because, during their evolution, plants never developed a membrane that was permeable to CO2 but impervious to water vapour. For the exchange of these two gases there are special openings in the leaf epidermis called stomates. The contra-flow gas exchange of water vapour and CO2 that takes place between the inside of a leaf and the atmosphere through the stomates is therefore unavoidable. If the exchange of gases is to be maintained for the production of dry matter, growth and development, plants require a continual supply of water in liquid form. This is particularly true for plants other than succulents and halophytes, since the internal store of water is normally very small relative to the daily loss. The steady use of water demands its continual uptake.
There is another reason why plant life is immediately dependent on water. In contrast with animals, land plants live permanently in one place, so they have to remove water from the soil water reservoir in their immediate vicinity. Plant life depends essentially on water that is stored within the soil and is available for extraction. For extraction of water, plants rely on their root systems, which continue to grow through most of their life. The quality of the soil as a store of water accessible to roots depends on texture and structure.
The daily throughput of water, that is the removal of water from the soil by roots, its movement through the plant in liquid phase, and its final transfer to the atmosphere in the vapour phase, can amount to a considerable quantity in comparison with the mass of the plants involved. In the middle of June, 1 week before heading, the dry weight of an oat crop amounted to 400 g m–2 (4 t ha–1). The daily water use at this stage of development was equivalent to 6 mm of precipitation, which in this case came from the soil storage (Ehlers et al., 1980a). These 6 mm of water throughput convert into 6 l or about 6 kg of water m–2 or 60,000 l (approximately 60 t) ha–1. Hence, relative to the dry mass of the standing crop, 15 times more water was returned to the atmosphere on a daily basis. Assuming that 85% of the shoot mass was water, the oat crop contained 2270 g water m–2. Compared with this store of water in the shoot, 2.6 times more water was extracted from the soil and passed on to the atmosphere. This transfer of water by plants to the atmosphere in the form of vapour is called transpiration.
Box 1.1. Light and water – prerequisites of photosynthesis
In the so-called light reaction of photosynthesis water is split into oxygen, protons and electrons:
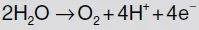
The oxygen produced moves out of the plant by diffusion preferentially through the stomates. At the same time during this light reaction nicotinamide adenine dinucleotide phosphate (NADP) is reduced to NADPH and in a coupled process adenosine diphosphate (ADP) is phosphorylated by use of inorganic phosphate (Pi), forming the ‘energy-rich’ adenosine triphosphate (ATP). NADPH and ATP as well as enzymes bring about the fixation of CO2 in the so-called dark reaction. In this reaction CO2 is reduced, ATP is dephosphorylated and NADPH is oxidized. The CO2 gets assimilated, and organic compounds can then be built (Fig. B1.1).
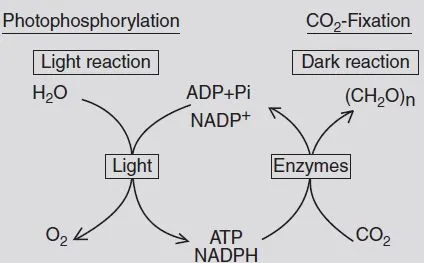
Fig. B1.1. Light and dark reactions during photosynthesis (after Gardner et al., 1985).
The enzyme involved in the primary process of CO2 assimilation is known as ribulose-1,5-bisphosphate (RuBP) carboxylase. That is true for the C3 plants (see Section 1.2), but for the C4 plants it is another enzyme, named phosphoenolpyruvate (PEP) carboxylase. The latter enzyme is also involved in CO2 assimilation by certain succulent plants. These plants have the capability of crassulacean acid metabolism (CAM).
Clearly we can say that the demands of plants cannot be satisfied with a small amount of water. Certainly it can be said that nature allows plants to be prodigal with this resource. The amount of water that is transpired daily by plants is generally 1–10 times more than the water stored in them. Compared with the amount needed for cell division and cell enlargement, the amount is 10–100 times more, and finally compared to the needs for photosynthesis it is 100–1000 times greater. So that prompts the question: ‘Why is all the water taken up?’
Water is an important constituent of all plants. Roots, stems and leaves of herbaceous plants are 70–95% water. In contrast, water comprises only 50% of ligneous tissues and 5–15% of dormant seeds.
Water is the basis of life for a single cell and for the aggregate of cells that combine to form the structure of higher plants. It not only influences the processes and activities of cell organelles, but can also determine the final appearance of a plant. As a chemical agent it takes part in many biochemical reactions, for instance in assimilation (Box 1.1) and respiration. It is a solvent for salts and complex molecules, and mediates chemical reactions. Water is the medium of transport for nutrient elements and organic molecules from the soil to the root and the means of transport of salts and assimilates within the plant. Stimulation and motion of organelles and cell structures, cell division and elongation are examples of processes controlled by hormones and growth substances, with water being the carrier of these messengers, enabling the regulatory system of the plant.
Other functions of water are much more apparent. Water confers shape and solidity to plant tissues. If a previously sufficient supply of water is disrupted, herbaceous plants and plant organs that lack supporting sclerenchyma will lose their strength and wilt. The hydrostatic pressure in cells is dependent on their water content, and permits cell enlargement against pressure from outside, which originates either from the tension of the surrounding tissue or from the surrounding soil. Root tips experience a confining pressure when penetrating soil because the particles, held together by cohesive and adhesive forces, have to be pushed apart to allow the root to penetrate. The large heat capacity of water greatly dampens the daily fluctuations in temperature that a plant leaf might otherwise undergo, due to the considerable amount of energy required to raise the temperature of water. Energy is also required to convert liquid water to a vapour that transpires from leaves causing cooling due to evaporation. Without these temperature compensating effects, plants would warm up much more and eventually die from overheating. Interestingly, because of these effects, transpiration rates can be estimated from surface temperatures, obtained by infrared thermography, either immediately within the crop stand (Section 14.1) or using remote sensing from aeroplanes or satellites.
1.2 Adaptation Strategies of Plants to Overcome Water Shortage
Depending on the amount and distribution of rainfall and the probability of occurrence, the regions of the world vary greatly in the supply of water. The support to plant life ranges from great abundance to extreme poverty. Plants have developed various strategies to counter the problems of temporal or spatial water shortage.
According to the presence and supply of water, ecologists divide terrestrial plants into hygrophytes, mesophytes and xerophytes. Hygrophytes are plants that thrive in generally humid habitats, where there is no shortage of water throughout the growing season. In temperate zones, in addition to these plants with a humid biotype, there are many shade-loving herbaceous forest species that also belong in this category.
At the opposite end of the spectrum are the xerophytes. These plants are adapted to water shortage, which may occur regularly and may persist over long periods of time. Anatomical and physiological specialization has taken place to meet the requirements of these plants so that they can survive extended periods of water scarcity. To this group belong succulent plants that establish an internal water reservoir for use during dry spells, thereby postponing desiccation. Another group of xerophytic plants are able to endure considerable water loss from their tissues without losing their ability to survive.
Mesophytes fit in between these two extremes. Many plants from temperate climates belong to this group, but the cultivated plants from those regions are also included. The latter cannot endure an extreme form of arid climate without being irrigated. However, they are well prepared for short periods of water shortage. When water supply falls short, they can reduce their transpiration rate dramatically and modify other processes.
Weather patterns may result in temporal and spatial shortages in water supply with varying intensity. As mentioned earlier, rainfall is a principal factor responsible for limitations in the plant water supply. But water shortages depend also on a second climatic var...
Table of contents
- Cover
- Half Title
- Dedication
- Title
- Copyright
- Contents
- Preface to 2nd Edition
- Preface to 1st Edition
- Abbreviations
- 1 The Role of Water in Plant Life
- 2 The Role of Water in Soil
- 3 The Interdependence of Soil Water and Vegetation
- 4 Properties and Energy State of Water
- 5 Water Storage and Movement in Soil
- 6 The Root – the Plant’s Organ for Water Uptake
- 7 The Water Balance of the Plant
- 8 The Plant as a Link between Soil and Atmosphere: an Overview
- 9 Water Use by Crops
- 10 Radiation and Dry Matter Production
- 11 Water Use and Dry Matter Production
- 12 Influence of Nutrient Supply on Water Use and Establishment of Yield
- 13 Development of Economic Yield under Inadequate Water Supply
- 14 Water Stress in Plants
- 15 Climatic Factors Influencing Yield
- 16 Breeding for Yield and Water Use
- 17 Controlling the Soil’s Water Balance by Soil Management
- 18 Controlling Water Use by Crop Management
- 19 Irrigation
- 20 Epilogue
- References
- Index
- Back
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Water Dynamics in Plant Production by Wilfried Ehlers,Michael Goss in PDF and/or ePUB format, as well as other popular books in Technology & Engineering & Environmental Management. We have over 1.5 million books available in our catalogue for you to explore.