Salmonella remains a major cause of economic loss in domestic livestock and human food poisoning worldwide. In the last 10 years there have been major advances in understanding the salmonella organism, meaning a compiled source of the new research is urgently needed. With fully updated chapters and new coverage of genome structure, virulence, vaccine development, molecular methods for epidemiology and exotics, this second edition is an invaluable resource for researchers of animal and human health.

- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Salmonella in Domestic Animals
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
1 Taxonomy and Species Concepts in the Genus Salmonella
‘When I use a word,’ Humpty Dumpty said, in rather a scornful tone, ‘it means just what I choose it to mean – neither more nor less.’
‘The question is,’ said Alice, ‘whether you can make words mean so many different things.’
‘The question is,’ said Humpty Dumpty, ‘which is to be master – that’s all.’
From Through the Looking Glass, Lewis Carroll
Taxonomy of Bacteria
The word ‘species’ has been used by scientists for many years; different people use it with different meanings, and in some cases individuals may use it with different meanings when they use it for different organisms. As in Humpty Dumpty, there have been many arguments over whether it is possible to be master of the word. The use of the word has been especially controversial in bacteria. In this review, we will see how the word species has been used in Salmonella taxonomy.
Humans have an inherent desire to classify objects in the world around them, in order to make sense of the apparently chaotic diversity that surrounds us. A small child, even at the age of a year or so, will obsessively sort objects such as marbles into piles according to size and colour. Biologists apply the same obsession to sorting the astonishing diversity of the living world into units called ‘taxa’; the hope is that this will help us to understand that diversity. This classification of the living world, though begun by others such as Aristotle, was codified and extended by C. Linnaeus in the 1600s. According to Linnaeus, the living world can be classified into species that are monotypic, immutable and that breed true (Rosselló-Mora and Amann, 2001). The original system of Linnaeus had five ranks: species were divided into subspecies, and the species were grouped into higher ranks of organisms including genera, orders and classes.
Though Darwin used the term in the title On the Origin of Species, he recognized that it had a very different meaning when used by different people at different times, for he wrote ‘I look at the term species as one arbitrarily given for the sake of convenience to a set of individuals closely resembling each other, and that it does not essentially differ from the term variety’ (Darwin, 1859). Taxonomists may take up the view of monism or pluralism; species monists believe that one species concept is correct for all organisms, while pluralists believe that different concepts are correct for different organisms (Doolittle and Zhaxybayeva, 2009; Ereshefsky, 2010a). Clearly, the species in prokaryotes is not the same as that of eukaryotes, so some form of pluralism must be applied.
Yet there are good arguments for maintaining the use of the term ‘species’ in bacteria as well as in other organisms, as summarized by Doolittle and Zhaxybayeva (2009) and by Ereshefsky (2010a, b). The use of species names is important in bacteria in many fields, including agriculture, biotechnology, food safety, disease diagnosis, epidemiology and public health and bioterrorism. It is desirable to attempt to apply the species name in a consistent and defendable way. But even where there is general agreement about the taxa and their relationships, there may be disagreement on which strains should be given the rank of species and thus how many species should be named. Perhaps the most dramatic differences between taxonomists who are called ‘splitters’ (who separate taxa into many species) and those who are ‘lumpers’ (who designate a small number of separate species) are found in taxonomists for Salmonella, where the splitters consider each serotype to be a separate species, so there would now be over 2600 species in Salmonella (Kauffmann, 1961), while the lumpers, according to the scheme now widely used, designate only two species, Salmonella enterica and S. bongori (Le Minor and Popoff, 1987; Reeves et al., 1989).
For those interested in understanding the diversity of Salmonella, determining and naming species is only part of the problem, because even within the serovar there is a startling variety of phage types, biotypes, electrophoretic types, sequence types, haplotypes etc., which represent real diversity in the population; this diversity must be understood in order to approach epidemiology in an informed way.
It is argued that successive speciation events created the tree of life; this is the notion that organismal diversity can be organized as ‘groups within groups’, which are formed as lineages successively separate, which yields a tree-like structure. However, this tree of life, first described by Darwin (1859), may not apply to bacteria where lateral gene transfer is an important mode of variation, so strains may have two quite distinct parents (Doolittle and Zhaxybayeva, 2009).
Introduction to Salmonella
Salmonella belongs to the family Enterobacteriaceae, and is a medically important pathogen for both humans and animals. Its main niche is the intestinal tract of humans and animals (Le Minor, 1991).
Salmonellosis in humans and animals can be broadly classified into two groups: enteric fever (typhoidal) and non-typhoidal salmonellosis (NTS) (Langridge et al., 2008). The disease can manifest itself in five ways: enteric fever, gastroenteritis, bacteraemia, extra-intestinal focal infection and a carrier state (Langridge et al., 2008) (Fig. 1.1).
The Kauffman–White scheme, first published in 1929, currently divides Salmonella into more than 2600 serotypes based on their serological reactions to somatic lipopolysaccharide (O), flagellar (H) and capsular (Vi) antigens (Grimont and Weill, 2007). However, these Salmonella serotypes can also be subdivided into the three following host range types that also influence clinical manifestations/presentations (Uzzau et al., 2000; Stevens et al., 2009): (i) host generalist (ubiquitous serovars) include Salmonella enterica serovar Typhimurium (S. Typhimurium) and S. Enteritidis that cause infections in diverse human and animal hosts; they produce a range of clinical symptoms but mainly acute and self-limiting gastroenteritis (Fig. 1.1) (Le Minor, 1991; Langridge et al., 2008; Stevens et al., 2009); (ii) host-adapted Salmonella (confined to a small number of hosts) such as S. Dublin in bovine animals and S. Choleraesuis in swine results in systemic infections, but these serovars can also infect humans and a limited number of other species (Le Minor, 1991; Langridge et al., 2008; Stevens et al., 2009); (iii) host-restricted Salmonella are associated with severe systemic infections in a single host: S. Typhi and S. Paratyphi cause enteric fever exclusively in man while S. Typhisuis causes paratyphoid in swine, S. Gallinarum causes typhoid in fowl and S. Abortusovis causes abortions in sheep (Le Minor, 1991; Stevens et al., 2009) (Fig.1.1).
Like many other infectious diseases, the course and outcome of the infection depends on variable factors including the dose of inoculation, the immune status of the host (age, underlying disease) and the genetic background of both the host and the infecting pathogen (e.g. virulence factors in the pathogen).
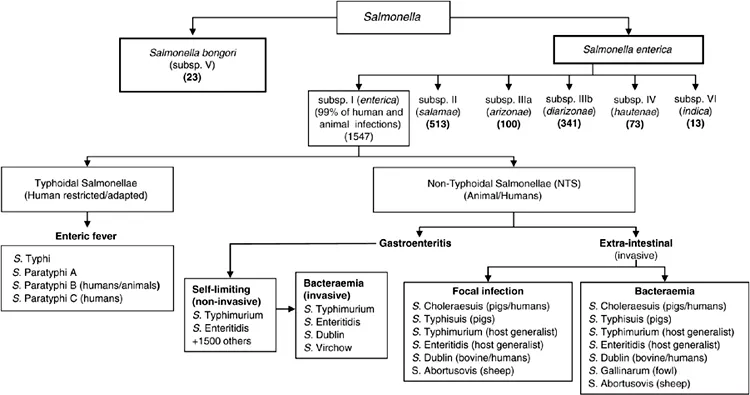
Fig. 1.1. Current scheme for classifying the genus Salmonella. Crosa et al. (1973) defined Salmonella to contain a single species, using DNA–DNA hybridization. A second species, designated S. bongori, was proposed by Reeves et al. (1989), using MLEE. Salmonella subspecies are defined by biotyping, DNA hybridization (Crosa et al., 1973), 16sRNA analysis and MLEE (Reeves et al., 1989). Serotyping is used to define serovars/serotypes within each subspecies. Numbers in brackets indicate the total number of serotypes included in each subspecies (Guibourdenche et al., 2010). Salmonella enterica subsp. I is adapted to endotherms. Sub species II is rarely found in humans but is commonly found in reptiles/ectotherms (Editorial team et al., 2010). Subspecies IIIa and IIIb are naturally found in reptiles but human infections have increased in recent years due to reptiles being kept as pets (Schröter et al., 2004; Editorial team et al., 2010). Serotypes of subsp. IV and VI are rare and mainly found in reptiles; they seldom cause infections in humans (Editorial team et al., 2010). Salmonella bongori is associated with ectotherms but has recently been isolated from endotherms (humans, birds) in Italy (Foti et al., 2009). Common serotypes are listed, but other serotypes may cause bacteraemia or focal infections. Reproduced from Langridge, G.C., et al., 2008. Invasive Salmonellosis in humans. Chapter 8.6.2.2 (revised version). In EcoSal – Escherichia Coli and Salmonella: Cellular and Molecular Biology. Böck, A., et al., (eds). ASM Press, Washington, D.C., Figure 1.1. with permission from the American Society for Microbiology.
Species Concepts
We will consider, in our discussion of bacteria in general and of Salmonella in particular, three of the (several) species concepts that have been discussed in eukaryotes and also in bacteria (Doolittle and Zhaxybayeva, 2009; Ereshefsky, 2010a).
The ecotype species concept (ESC) (which yields the eco-species) was described by Cohan (2001); each cluster of strains corresponds to an ecotype, defined as a population of cells in the same ecological niche. The concept has roots in the history of microbiology, since bacteria were initially considered to be asexual clones; something resembling this concept was used to determine species in Salmonella starting in the 1880s, when species were named according to the disease they caused and the host they infected. The ecotype theory invokes periodic selection within the variants that arise by mutation in an asexual clone; the mutant that is selected will out-compete others in the population, and the genomes carrying the mutant will sweep to fixation along with the selected mutant (Cohan, 2001). Thus other diversity, that was neutral in the clone’s members, will be purged. In this way, periodic selection limits variation in the genome and results in a population of cells that share a set of genes but are distinct from the population from which they were derived. Thus mutation (to produce variability) and selection both operate, but recombination has no role. In Cohan’s view, the bacterial species as presently recognized are analogous to eukaryotic genera, rather than to eukaryotic species.
The second is the biological species concept (BSC) (which yields the bio-species), a concept developed by Ernst Mayr (1996) based on observations in higher animals. Since members of the bio-species reproduce by sexual mating and mate only (or preferentially) with members of the same species, they share a gene pool and the species maintains genetic cohesion. Bacteria were initially thought to be asexual; however, genetic recombination has been recognized in bacteria since the 1940s and 1950s (Lederberg and Tatum, 1946), but there are three differences between recombination in bacteria compared with those eukaryotes that reproduce sexually: reproduction in bacteria is due to binary fission, while sexual exchange if it occurs is (usually) rare and not linked to reproduction; there is only partial transfer of the genome of one parent, rather than fusion of gametes from both parents as in eukaryotes; genetic transfer in bacteria is unidirectional, with transfer from the donor to the recipient.
The total process of recombination in bacteria takes place in two steps; the first step is genetic transfer from donor to recipient to form a partial diploid zygote. The donor contribution may be a few kilobases in DNA-mediated transformation, or a few tens of kilobases in phage-mediated transduction, or up to hundreds of kilobases in plasmid-mediated conjugation (Snyder and Champness, 1997). The second step in the process is integration of the donor DNA into the recipient chromosome to restore the haploid state. (In some cases an intact replicon such as a plasmid is transferred, which does not have to integrate, but these cases are not further considered here.) Integration may occur by two different methods.
The first method is homologous recombination, which requires the RecA protein and occurs between regions of DNA in the donor and recipient that have a high degree of sequence identity: the allele from the donor replaces the allele of the recipient in a process called allelic replacement (Snyder and Champness, 1997). Thus, allelic replacement can insert genes from closely related bacteria but not from unrelated bacteria. Allelic replacement has the theoretical potential to form a gene pool with resulting genetic cohesion; members of this gene pool might form bio-species. We will see below that allelic replacement does occur in S. enterica.
The second method is site-specific recombination, which can insert fragments of DNA from the donor into the recipient chromosome by mechanisms that are not dependent on RecA protein and do not require DNA with high sequence identity (Nash, 1997). These site-specific mechanisms include insertion of prophages, transposons, or other DNA units. This results in lateral gene transfer (LGT) (also called horizontal gene transfer); it adds non-homologous blocks of DNA to the genome of a bacterium, in sizes ranging from a few kilobases to hundreds of kilobases. This process, like mutation, contributes to variation in the population, not to genetic cohesion as in allelic replacement, and therefore it would not contribute to forming bio-species. The consequence of LGT is that the genome of bacteria is composed of two types of genes: the core genome, which includes genes that are common to all the members of the group being studied, and the accessory genome, which includes genes that are missing from one or more of the members of the group being studied and which have resulted from LGT (Lan and Reeves, 2001). Thus, 89% of the genes of two serovars of Salmonella, Typhimurium strain LT2 (McClelland et al., 2001) and Typhi strain CT18 (Parkhill et al., 2001), are orthologues with nucleotide sequence identity usually higher than 99% (the core genome), while 11% of the genes do not have orthologues in the other member of the pair of serovars (the accessory genome). The core genome is not a fixed entity, but is determined by the strains being compared; as the number of strains in the comparison increases, the size of the core genome reduces and the size of the accessory genome increases. If all sequenced bacteria are included, the size of the core reduces to only about 20 genes, including only those involved in central processes such as replication and protein translation (Bapteste et al., 2009). Taxonomic analysis usually studies the core genome, using sequence-based methods (such as multi-locus sequence typing, described below), though it can also study the accessory genome, using methods such as microarrays (Porwollik et al., 2002).
The third concept is the phylogenetic species concept (PSC) that yields the phylospecies, in which the lineage of the cells is inferred from the degree of identity of nucleotide sequences. Strains that show >70% relatedness according to DNA–DNA hybridization (DDH) carried out under rigorously controlled conditions are placed in the same species according to many authorities including Staley (2006), though Crosa et al. (1973) consider >70% relatedness to indicate belonging to the genus Salmonella, and >80% belonging to the same species. Strains that have >97% identity in sequence of 16S rRNA are placed in the same s...
Table of contents
- Cover Page
- Title Page
- Copyright Page
- Contents
- Contributors
- Preface
- 1 Taxonomy and Species Concepts in the Genus Salmonella
- 2 Structure, Function and Synthesis of Surface Polysaccharides in Salmonella
- 3 Fimbriae and Flagella of Salmonella enterica
- 4 Characterizing Salmonella Genomes
- 5 Salmonella Virulence Mechanisms and their Genetic Basis
- 6 Host Genetic Susceptibility/Resistance
- 7 Antimicrobial Resistance in Salmonella
- 8 Immunity to Salmonella in Farm Animals and Murine Models of Disease
- 9 Salmonella Infections in the Domestic Fowl
- 10 Salmonella Infections in Turkeys
- 11 Salmonella Infections in Ducks
- 12 Salmonella Infections in Cattle
- 13 Salmonella Infections in Pigs
- 14 Salmonella Infections in Sheep
- 15 Salmonella Infections in Horses
- 16 Salmonella Infections in Dogs and Cats
- 17 Salmonella Infections in Exotic Pets
- 18 Public Health Aspects of Salmonella Infections
- 19 Salmonella in Feed
- 20 Environmental Aspects of Salmonella
- 21 Competitive Exclusion
- 22 Vaccination against Salmonella Infections in Food Animals: Rationale, Theoretical Basis and Practical Application
- 23 EU Legislation on the Control of Salmonella, Monitoring and Reporting
- 24 Current and New Approaches to Typing of Salmonella
- 25 Other Approaches to Infection Control
- Index
- Footnotes
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Salmonella in Domestic Animals by Paul Barrow, Ulrich Methner, Paul Barrow,Ulrich Methner in PDF and/or ePUB format, as well as other popular books in Medicine & Veterinary Medicine. We have over 1.5 million books available in our catalogue for you to explore.