![]()
Chapter 1
How Biology Solved Its Energy Problem and Implications for the Future of Humankind
James Barber
The appearance of life on our planet occurred about four billion years ago following five hundred million years of geological and chemical evolution. It took the form of single cell prokaryotic chemotrophs obtaining their energy from chemicals which could act as electron donors. After 1.5 billion years there were a wide variety of anaerobic prokaryotes which fell into two groups, archaea and bacteria. Darwinian principles had allowed these prokaryotes to be widely distributed in many different types of environments and evolve complex systems to capture and utilise energy for the reductive process of cellular biochemistry. Sooner or later, however, the energy sources (oxidisable substrates) available to these chemotrophic prokaryotes became limiting and some developed the capacity to supplement their metabolic energy requirements by absorbing solar radiation. This heralded the “Big Bang of Evolution” since this strategy gave rise to phototrophic organisms which could use sunlight to totally drive their bioenergetics. This was achieved by using solar energy to extract “hydrogen” (in the form of high energy electrons and protons) from water and release oxygen into the atmosphere. From then on, life prospered and diversified on an enormous scale driven by Darwinian evolution. It was from this success that humans evolved as a very special species with abilities far greater than any other animal that has ever lived on planet Earth. Yet we are on a very dangerous course which some believe could result in the elimination of our species in a matter of a few hundred years if we carry on the way we do today, that is, continue to use fossil fuel as a major energy source. Biology solved its energy problem by using sunlight to split water and extract the stored energy by recombining the “hydrogen” with oxygen via respiration. We must do the same by using energy from the sun captured directly via photovoltaic cells and indirectly by hydro-, wind- and wave-power to give electricity and hydrogen as the two main energy carriers. The latter complements batteries for renewable energy storage and must come from the electrolysis of water. There is an urgent need to progress with expanding and developing the appropriate technologies and putting in place the infrastructure, particularly for the generation of hydrogen and its storage, transport and use.
1.Evolution of Bioenergetics
1.1.Under anaerobic conditions
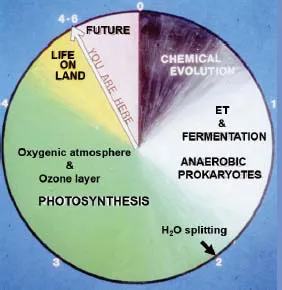
According to the Big Bang Theory, the Universe came into being about 13.7 billion years ago, leading to the formation of our Planet and Solar System about 9.2 billion years later. As depicted in Figure 1, it took half a billion years for the Earth to cool and establish a chemical environment for life to evolve. Primordial Earth must have been a very hostile environment. Just how the complex reactions that define life came about remains a mystery. It is usually assumed that it took place in a soup of organic and inorganic molecules in a very specific aqueous environment [Sarchet, 2018], possibly around a hydrothermal vent [Sojo et al., 2016] or in a shallow pond exposed to intense UV light [Mulkidjanian et al., 2012].
Present day knowledge of molecular genetics and cell biology indicates it was a unique event and all life can be traced back to a single starting point leading to the Last Universal Common Ancestor (LUCA). Within the primeval soup polymers of nucleic acids would need to form which could act as templates for replication. It is not so difficult to imagine this chemistry occurring given the properties of polynucleotides and rate of chemical mutations that could occur over very long periods of time. Ultimately these chains of nuclear acids could act as templates for amino acids also thought to have been present in the primeval soup. It seems almost incomprehensible that from these initial chemical events that life would evolve to the complexity of anaerobic prokaryotic organisms similar to those found on our planet today. However to rationalise this incredible achievement it has to be recognised that it occurred over an almost inconceivable period of time, in the region of about 1.5 billion years (Figure 1). For this reason alone we should respect and marvel at the evolution of life on our planet.
Figure 1.A five billion year clock of evolution of life on our planet. ET = electron transport.
The early forms of anaerobic life had to establish a myriad of enzymes necessary for the prokaryote cell to grow and reproduce and be able to adapt to different environments. But this massive achievement required a continuous input of energy which would have been supplied from the stock of high energy organic and inorganic molecules created during the period of chemical evolution. Sooner or later the reductive chemistry would have become more reliant on sources such H2, CO and CH4 as electron donors and evolve the ability to generate electrochemical potential gradients across the cell membrane and convert this energy source to adenosine triphosphate (ATP). This is rather like that found in oxygenic organisms but with no O2 available as a terminal electron acceptor [Olsen, 2006]. Instead they could use a range of electron acceptors such SO4, NO and S. As the biochemistry of these early stages of bioenergetics advanced, fermentation pathways must also have evolved to obtain ATP without the need for an electron transport chain. For all this to occur, a cell membrane had to exist which almost certainly started off as vesicles consisting of a double layer of amphiphilic lipids (liposome) which form readily when these molecules are agitated in an aquatic environment, for example by the bursting of bubbles in the water close to hydrothermal vents or at the water surface by wave action. The formation of these vesicles could encapsulate organic molecules and inorganic ions present in the primeval soup. At that time the CO2 level in the environment was high and several biochemical pathways seemed to emerge to aid its reduction [Apel et al., 2013]. Coupled with this would be the requirement to maintain osmotic stability so that energy driven pumping mechanisms across the cell membrane would have to be established. It seems much of this evolved during LUCA and thus before the diversion to archaea and bacteria around 3.8 billion years ago [Ouzounis et al., 2000; Weiss et al., 2016].
Under some circumstance an energy crisis almost certainly would have developed when chemically-based reductants were not sufficient. Amazingly some organisms developed a way to supplement their metabolism utilising the energy of sunlight. Certain archaea developed their own “photovoltaic cells” as represented today by Halobacteriaceae. When there is a shortage of chemical substrates they produce a pigmented cell membrane containing bacteriorhodopsin [Sharma et al., 2006]. This pigment undergoes a conformational change on absorbing light and in so doing, drives the accumulation of energised protons across the cell membrane which can supplement the metabolic energy requirement by powering the production of ATP. But far more important for future evolution of life was the appearance of anaerobic photosynthetic bacteria about 3.5 billion years ago [Olsen and Blankenship, 2005; Hohmann-Marriott and Blankenship, 2011]. In this case chlorophyll was the pigment of choice. Unlike bacteriorhodopsin, the absorption of light energy by bacteriochlorophyll results in charge separation with the electron elevated to a high energy level. The energy rich electrons could then be used to supplement the energy demands of these photosynthetic bacteria. The positive hole left in the chlorophyll is neutralised by an electron donor. The donors could now be less reducing and a range of organic and inorganic species were likely to be available including SO4, NO3, H2S, NH3, and various organic acids. Of particular importance would have been the availability of multivalent reduced metal ions as electron donors, particularly Fe2+, but of considerable significance would be the availability of Mn as an electron donor which would ultimately be the basis of the water splitting catalytic centre. Mn would have been very abundant and widely distributed as it is today and available as Mn2+ in solution or in a mineralised form [Sauer and Yachandra, 2002; Najafpour, 2011]. This heralded a major step in the evolution of life on our planet.
Before this occurred anaerobic prokaryotic life had been highly successful and Darwinian principles would have driven this success. It is reasonable to assume that those organisms which utilised light energy had a distinct advantage over those who could not. This would mean that the aquatic environment would be coloured. Goldsworthy [1987] has made the point that the absorption spectrum of bacteriorhodopsin is in the green part of the solar spectrum while that of bacteriochlorophyll is in the near UV/blue and far red/near IR regions. This would allow both types of organisms to live side-by-side. However halobacteria have the disadvantage of requiring a supply of energy rich substrates to drive their reductive metabolism while photosynthetic bacteria could exist on more abundant less reducing electron donors. On the other hand halobacteria are archaea rather than bacteria, many of which can grow in extreme environmental conditions and presumably in such cases survived where photosynthetic bacteria could not. This is the case today. Based on the existence of modern anaerobic photosynthetic bacteria we can assume there were two classes differing in their photosynthetic reaction centres (RC); with Type 1 RC where the terminal electron acceptors are ferredoxins (e.g. heliobacteria, green sulphur bacteria, green non-sulphur bacteria) and Type II RC where quinones act as terminal electron acceptors (e.g. purple bacteria such as Rhodobacter sphaeroides, Rhodopseudomonas viridis). Both types of RCs consist of bacteriochlorophyll absorbing in the near far red (850 nm to 960 nm) and they share structural features which indicate that they are derived from a common ancestor.
It is often said that before the water splitting mechanism evolved, anaerobic prokaryotes were simple forms of life. This is far from the truth. They were already very advanced in their cellular metabolism and bioenergetics much of which was established during LUCA and therefore before the separation into archaea and bacteria. They were able to adapt to many different, and often extreme environmental niches. But inevitably their future development was limited by the resources available to them, particularly energy. However the complex machinery of anoxygenic bacterial photosynthesis and the associated respiratory complexes had evolved and established a framework for aerobic bioenergetics.
1.2.Under aerobic conditions
Clearly having the ability to use light energy to supplement or fully drive cellular metabolism has a distinct evolutionary advantage [Nelson, 2011]. Even so, ultimately the biomass of anaerobic prokaryotes would be limited by the availability of appropriate electron donors. As emphasised above, this energy problem was solved when organisms evolved the ability to split water using sunlight. Using water as an electron donor, coupled with the production of O2 as a by-product, set the scene for the “Big Bang of Evolution.” The release of this gas about 2.6 billion years ago had dramatic consequences for the development of life, since it created an oxygenic atmosphere and in time allowed the ozone layer to form [Canfield et al., 2005; Fischer et al., 2016]. With oxygen available, the efficiency of cellular metabolism increased dramatically, since for a given amount of substrate, aerobic respiration provides in the region of 20 times more energy than anaerobic respiration. It was probably this improved efficiency of cellular bioenergetics which paved the way for the subsequent evolution of eukaryotic cells and multicellular organisms. The establishment of the ozone layer provided a shield against harmful UV radiation allowing organisms to exploit the terrestrial, as well as the aquatic environments. Consequently life has been able to prosper and diversify on an enormous scale as testified by the fossil records and by the extent and variety of living organisms on our planet today. For this to happen several major steps had to be made:
(i)Incorporation and electronic coupling of Type 1 and Type 2 RCs into the same organism so that the energy of two photons of light could span the energy gap between the oxidation of water and the reduction of ferredoxin and also power ATP formation (photophosphorylation).
(ii)Modification of the synthesis of chlorophyll so that its spectral properties did not compete with those of bacteriochlorophyll and also be able to use a more energy rich part of the lon...