Environmental Experience and Plasticity of the Developing Brain goes beyond the genetic basis of neurodevelopment. Chapters illuminate the external factors that can dramatically impact the brain early in life and, consequently, the eventual accomplishment of developmental milestones and the construction of adult behavior and personality.
Authored and edited by leaders in this rapidly growing field, Environmental Experience and Plasticity of the Developing Brain not only surveys preexisting literature on the effects of environment versus genetics, but also discusses more recent studies on the impacts of neurodevelopment in terms of maternal stimulation, environmental enrichment and sensory deprivation. The book also includes key examples of environmental impacts on preexisting genetic syndromes leading to developmental disabilities. Focus is also given to the consequences of early adverse experience in primates, as well as neurobiological and behavioral consequences in institutionalized human children and the reversibility of such consequences.
Environmental Experience and Plasticity of the Developing Brain encompasses a broad area of research in the field of developmental neurobiology and offers a unique combination of different examples of environmental factors affecting brain development and behavior.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Chapter 1 Environmental enrichment and brain development
Alessandro Sale1, Nicoletta Berardi2 and Lamberto Maffei1
1Neuroscience Institute, National Research Council (CNR), Pisa, Italy
2Department of Neuroscience, Psychology, Drug Research and Child Health NEUROFARBA, University of Florence, Florence, Italy
Introduction: critical periods and experience-dependent plasticity in brain circuits
The term “plasticity” refers to the ability of the nervous system to reorganize its connections functionally and structurally in response to changes in environmental experience. This property underlies the adaptive development and remodeling of neuronal circuitry that makes brain development, behavioral flexibility, and long-term memory possible.
Plasticity is particularly high during developmental time windows called critical periods (CPs), when experience is crucial in promoting and regulating neural maturation and, consequently, the behavioral traits of the newborn, in every vertebrate species tested so far, from birds to rodents to primates (Berardi et al., 2000). Essentially, a CP is a phase of exceptionally high sensitivity to experience displayed by developing neural circuits. During CPs, experience exerts a key role in building the precise assembly of connections that endows each individual with his/her unique characteristics. Different species show different CPs for the same function, in good accordance with a different time course of development and life span. On the other hand, distinct functions show different CPs in the same species, correlating with different time courses of development in different brain areas.
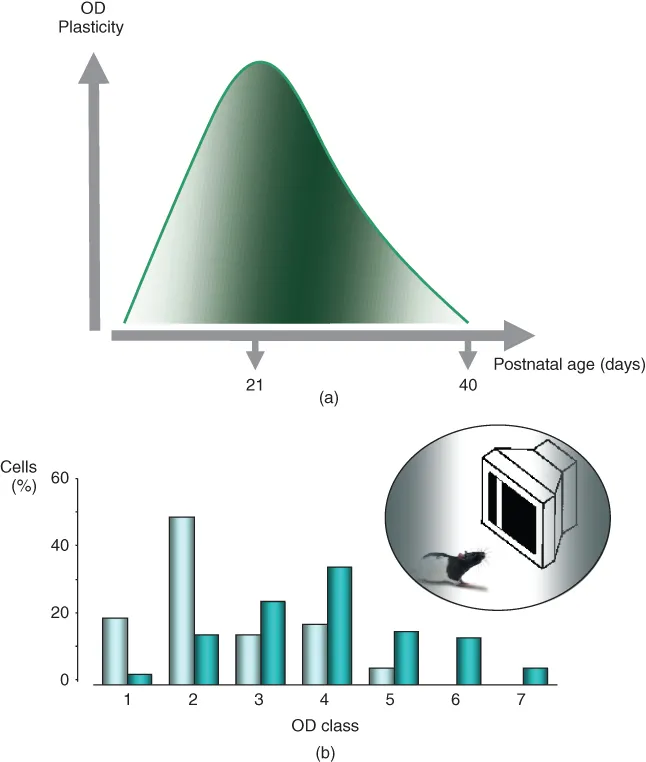
Essential information on developmental brain plasticity and CPs has been provided by studies focusing on the primary visual cortex (V1), which has been for decades the election model for studying experience-dependent plasticity in the brain. The pioneering experiments performed by Hubel and Wiesel showed how dramatically can early sensory deprivation affect the anatomy and physiology of the visual cortex (Figure 1.1). Many neurons in the visual cortex are binocular, that is, receive input from both eyes, and exhibit different degrees of dominance from either eye, a property called ocular dominance. Hubel and Wiesel reported that, early in development, reducing the visual input to one eye by means of lid suture, a treatment classically referred to as monocular deprivation (MD), disrupts ocular dominance of V1 cells, with a loss of neurons driven by the deprived eye and a strong increment in the number of cells driven by the open eye, and reduces the number of binocular neurons (Wiesel and Hubel, 1963). The imbalance of activity between the two eyes results in remarkable anatomical changes in V1, with a shrinkage of the deprived eye ocular dominance columns, those layer IV regions that receive thalamic inputs driven by the closed eye, and in the expansion of the open eye's columns (Hubel et al., 1977; Shatz and Stryker, 1978; LeVay et al., 1980; Antonini and Stryker, 1993), accompanied by a remodeling of cortical horizontal connections (Trachtenberg and Stryker, 2001). At the behavioral level, if the condition of MD is protracted for a long period during development, it eventually leads to lower than normal visual acuity and contrast sensitivity values for the deprived eye (amblyopia), together with a deterioration of binocular vision. Strikingly, the same manipulation of visual experience appeared to be ineffective in the adult (LeVay et al., 1980), leading to the characterization of the first and most widely studied example of CP (Berardi et al., 2000; Berardi et al., 2003; Knudsen, 2004; Hensch, 2005a, 2005b; Levelt and Hubener, 2012).
Figure 1.1 Critical period (CP) for ocular dominance plasticity in the rat visual cortex. (a) Schematic representation of the time course of CP for ocular dominance plasticity in the rat, which peaks around postnatal day (P) 21 and is definitively closed by the age of P45. (b) Single unit recordings from the primary visual cortex allow classification of neurons with respect to their ocular preferences: in a typical recording from a nondeprived animal (light cyan columns), cells in class 1 are activated exclusively by the contralateral eye, cells in class 7 are activated exclusively by the ipsilateral eye, neurons in classes 2–3 and 5–6 are activated to varying degrees by both eyes, and neurons in class 4 respond equally to both eyes. Following closure of the contralateral eye from 1 week during the CP, cells become much more responsive toward the ipsilateral open eye, at the expense of the deprived eye (dark cyan columns). (See insert for color representation of this figure.)
Another well-studied CP is that regulating age-dependent changes in fear memory acquisition, which in rodents emerges at the end of the second postnatal week of life (Akers et al., 2012). Interestingly, the potential for fear extinction does also follow a CP, displaying a permanent fear erasure in preadolescent mice but leading to incomplete erasure and thus persisting or returning fear responses in juveniles about 10 days older.
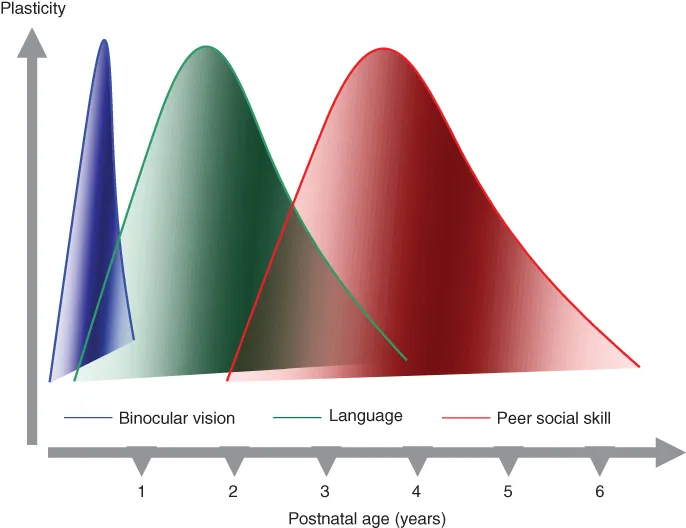
In humans, CPs have been documented for several brain modalities (Figure 1.2). Examples of CPs in the sensory domain are those for the maturation of visual acuity and stereopsis, the acquisition of language-specific abilities in phonemic perception, and the acquisition of gustatory and olfactory preferences (Lewis and Maurer, 2005; Werker and Tees, 2005; Ventura and Worobey, 2013). A particularly relevant case of olfactory learning regulated by a CP is that underlying maternal attachment, clearly present in newborn babies and well described at the neurobiological level in rodents (see also the chapter by Sullivan and colleagues in this book [Chapter 6]). CPs in humans have been also found for second language acquisition, both speech and sign language, or for proficient performance in musical instrument playing (Bengtsson et al., 2005; Kuhl, 2010).
Figure 1.2 Critical periods across brain functions in humans. The picture represents a schematic of the critical period time course for acquisition of binocular vision, language learning, and adequate peer social skills in children. Different functions display different time courses, both in terms of total duration of the heightened sensitivity window and concerning the age of onset and closure of the potential for plasticity. In the three curves, levels of plasticity have been normalized to the peak. (See insert for color representation of this figure.)
As in the case of MD, the importance of a proper experience during the CP is made particularly clear by the detrimental effects caused by its absence or deterioration, like in the classic example of the negative effects in the social/affective domain produced by rearing under conditions in which the mother is absent or early removed and sufficient maternal care levels are not available (Sullivan et al., 2006). Developmental plasticity, indeed, is by itself neither good nor bad, it simply takes its course, allowing the system to proceed toward an adaptive developmental trajectory when the stimuli are adequate and available, or instead resulting in severe and even permanent deficits under harsh environmental conditions. Thus, while the existence of a mechanism by which high levels of plasticity during the CP are followed by an abrupt reduction of circuit modifiability after its closure is likely to provide adaptive advantages in terms of the possibility to fix the acquired neural assemblies without the need of continuous maintenance, it may also expose the nervous system to severe dysfunctions when development is perturbed.
Importantly, the potential for recovery after reestablishment of proper environmental conditions can also be regulated by CPs, with studies in postinstitutionalized children demonstrating that the most severe and persisting effects of raising children in impoverished environments lacking sufficient social stimuli are more likely to be documented when adoption occurs beyond 4–6 months of age (for a comprehensive survey of the literature on the effects of institutional deprivation, see the chapter by Doom and Gunar in this book [Chapter 9]).
Not surprisingly, much effort in Neuroscience research is currently devoted to understanding the molecular mechanisms underlying the closure or the sudden reduction of plasticity at the end of the CPs. Among the most promising candidates are factors exerting a key role as plasticity brakes, such as critical components of the extracellular matrix, that is, the chondroitin sulphate proteoglycans that surround neuronal cell bodies in structures called perineuronal nets, myelin-related Nogo receptors, proteins belonging to the newly discovered class called Lynx family, epigenetic regulators of the functional state of chromatin such as histone deacetylase inhibitors, and the maturation of intracortical GABAergic interneurons (Bavelier et al., 2010; Nabel and Morishita, 2013).
Optimization of environmental stimulation: environmental enrichment
In parallel to the Hubel and Wiesel seminal work based on a sensory deprivation approach, fundamental contributions to the knowledge of how experience affects brain development have been provided by the group of Rosenzweig and colleagues, using the so-called environmental enrichment (EE) paradigm. Originally defined as “a combination of complex inanimate and social stimulation” (Rosenzweig et al., 1978), EE is performed in wide cages where the animals are reared in large social groups and in the presence of a variety of objects, like tunnels, nesting material, stairs, and plastic recoveries, that are changed by the experimenter at least once a week in order to stimulate the explorative behavior, curiosity, and attentional processes of the animals. An essential component of EE is voluntary physical exercise, the opportunity to attain high levels...
Table of contents
Cover
Dedication
Title Page
Copyright
Table of Contents
List of contributors
Chapter 1: Environmental enrichment and brain development
Chapter 2: Epigenetic control of visual cortex development and plasticity
Chapter 3: Gene–environment interactions in the etiology of psychiatric and neurodevelopmental disorders
Chapter 4: Critical periods and neurodevelopmental brain disorders
Chapter 5: Maternal care and DNA methylation
Chapter 6: Neurobiology and programming capacity of attachment learning to nurturing and abusive caregivers
Chapter 7: Early environmental manipulations and long-term effects on brain neurotrophin levels
Chapter 8: Effects of genes and early experience on the development of primate behavior and stress reactivity
Chapter 9: Institutional deprivation and neurobehavioral development in infancy
Chapter 10: Impact of infantile massage on brain development
Index
End User License Agreement
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Environmental Experience and Plasticity of the Developing Brain by Alessandro Sale in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Neuroscience. We have over 1.5 million books available in our catalogue for you to explore.