During the course of evolution, life on the Earth started in a reducing environment and about 3.2 billion years ago, the reducing environment changed to an oxidizing one due to the appearance and proliferation of the first oxygen‐evolving photosynthetic organisms, that is, cyanobacteria (Schopf
et al., 2007). In other words, the cyanobacteria are considered to be the first organisms to release oxygen in the environment by means of an oxygen evolving complex (OEC) (Bekker
et al., 2004). The outermost orbital of the dioxygen (O
2) molecule has two unpaired electrons having same spin quantum number, and this enables O
2 to accept electrons one at a time efficiently, and generate the reactive oxygen species (ROS). Out of the total O
2 utilized by plants, 1% is diverted to produce ROS in various cell organelles (del Rio
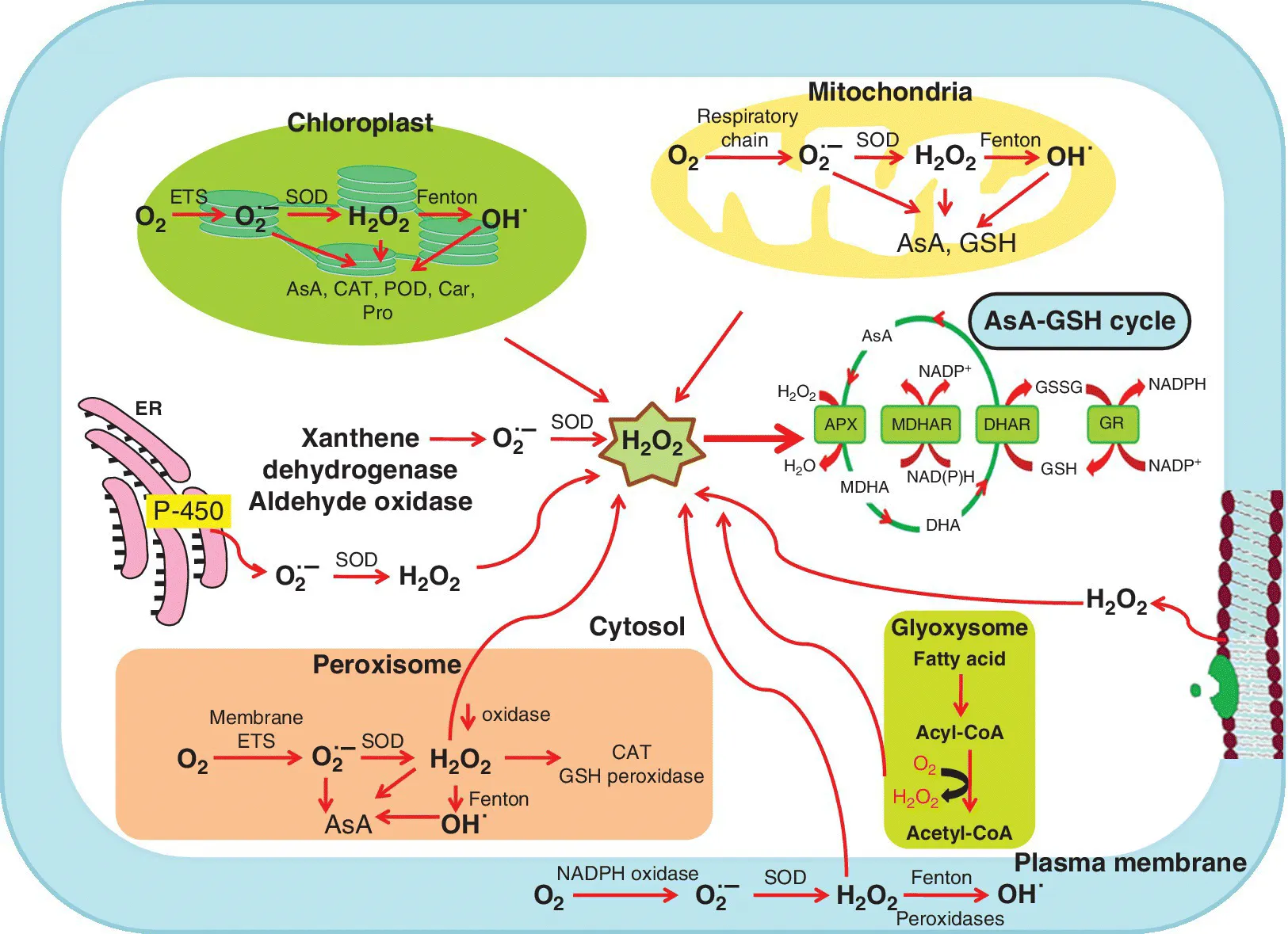
et al., 2002). Reactive oxygen species are essential by‐products of all aerobic organisms that are produced during normal metabolic processes as well as under stress conditions. The ROS‐producing subcellular organelles are mainly mitochondria, chloroplasts, peroxisomes, cytosol, and plasma membrane (Corpas
et al., 2015; Hasan
et al., 2016) (
Figure 1.1). The roles of ROS are contradictory, they may have negative as well as positive roles depending upon their concentrations in the particular cell organelles. At higher concentrations, ROS cause damaging effects on proteins, DNA/RNA, and lipids by oxidative modification in plant cells (Gill and Tuteja, 2010; Hasan
et al., 2016). On the other hand, previous evidence clearly showed that at lower concentrations ROS act as signaling molecules in plants for regulating developmental pathways and control of redox homeostasis and defense responses against pathogens and environmental stress (Wood
et al., 2003; Apel and Hirt, 2004). There are well‐described mechanisms in prokaryotes where the concentration of ROS directly activates transcription factors that overexpress the genes to combat oxidative stress (Kiley and Storz, 2004). There are several ROS, such as superoxide radical (
), hydroxyl radical (·OH), hydroperoxyl radical (HO
2·), hydrogen peroxide (H
2O
2), alkoxy radical (RO·), peroxy radical (ROO·), singlet oxygen (
1O
2), and excited carbonyl (RO*), all of which are cytotoxic to plants at elevated concentrations (Dismukes
et al., 2001; Karuppanapandian
et al., 2011). In the cell organelles accumulation of superoxide enhances oxidative stress rather than playing a role in redox signaling. However, in some cases it damages certain proteins that activate specific signaling pathways
and consequently leads to death of the particular cell (Chen
et al., 2009). Hydrogen peroxide (H
2O
2) acts as a signaling molecule that diffuses across membranes and triggers specific signal transduction pathways (Veal and Day, 2011). The balance between production and elimination of ROS is dependent upon various biotic and abiotic factors such as temperature, heavy metal concentration, drought, salinity, UV radiation, light, nutrient deficiency, and excessive use of pesticides and/or herbicides as well as pathogen attacks. This disturbance ultimately leads to
increased concentration of ROS in particular cell organelles. The damaging effects of ROS are ameliorated by different antioxidative defense systems. The antioxidant system consists of enzymatic antioxidants, namely superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and glutathione
S‐transferase (GST), ascorbate peroxidase (APX) and non‐enzymatic antioxidants, including non‐protein thiols (NP‐SH) ascorbate, tocopherol, caretenoid and cysteine. These act together as a machine to detoxify ROS (Kumar
et al., 2015; Singh
et al., 2016; Tripathi
et al., 2012a,b, 2016a,b) (see
Figure 1.1). Among these, SOD is considered as first line of defense; it dismutates the superoxide and subsequently H
2O
2 is generated. Further, H
2O
2 detoxification involves different enzyme systems in distinct cellular compartments. The peroxidase family includes ascorbate peroxidase (APX), glutathione peroxidase (GPX), and peroxidase (POD) for the elimination of H
2O
2. Another enzyme, CAT, is important in the removal of H
2O
2 generated in peroxisomes. GPX also reduces H
2O
2 as well as organic and lipid hydroperoxides by using glutathione (GSH) as substrate. Among the various H
2O
2‐detoxifying enzymes, APX plays the most essential role in scavenging ROS. APX is present in thylakoid, glyoxisome, chloroplast stroma, and cytosol, and is involved in the scavenging of H
2O
2 through water‐water and ascorbate‐glutathione (AsA‐GSH) cycles, utilizing AsA as the electron donor. One of the antioxidant enzymes, GST, participates in herbicide detoxification, hormone homeostasis, and regulation of apoptosis and also is involved in plant responses to biotic and abiotic stresses. Non‐enzymatic antioxidants include the major cellular redox buffers ascorbate and glutathione, as well as tocopherol, flavonoids, alkaloids, and carotenoids.