Neurophysiology of Neuroendocrine Neurons provides researchers and students with not only an understanding of neuroendocrine cell electrophysiology, but also an appreciation of how this model system affords access to virtually all parts of the neuron for detailed study - something unique compared to most types of neuron in the brain. Chapters range from those describing the rich history and current state of in vivo recordings, highlighting the precise relationship between the patterns of action potential discharge in these neurons and hormone release, to in vitro approaches where neuroendocrine neurons can be precisely identified and their membrane properties, morphology, and synaptic responses, directly examined.
• Written by a team of internationally renowned researchers, each chapter presents a succinct summary of the very latest developments in the field • Includes an evaluation of different experimental approaches, both in vivo and in vitro, and how the resulting data are interpreted • Both print and enhanced e-book versions are available • Illustrated in full colour throughout
This is the first volume in a new Series 'Masterclass in Neuroendocrinology' , a co- publication between Wiley and the INF (International Neuroendocrine Federation) that aims to illustrate highest standards and encourage the use of the latest technologies in basic and clinical research and hopes to provide inspiration for further exploration into the exciting field of neuroendocrinology.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
SECTION 1A Magnocellular Neuroendocrine Neurons: Properties and Control of Vasopressin and Oxytocin Neurons
CHAPTER 1 Electrophysiology of Magnocellular Neurons In Vivo
Gareth LengNancy Sabatier
Centre for Integrative Physiology, University of Edinburgh, Edinburgh, UK
1.1 Introduction
Neuroendocrinology is the study of things that matter: stress and appetite, metabolism, body rhythms, growth, and all aspects of reproduction from the reproductive cycle, through sexual behavior, pregnancy and parturition, to lactation and maternal behavior—things that matter for our health and happiness. However, neuroendocrine systems are also influential model systems for neuroscience generally, because of their unique value as “windows on the brain.” The products of neuroendocrine systems can be measured relatively easily, and their effects are, with wit and persistence, determinable. In consequence, the neuronal activity that gives rise to those products is interpretable to a degree that can only be envied by colleagues in most other areas of neuroscience. If we ask of any neuron in the brain, what does it really do, the answers are often frustratingly incomplete: even if we know how it responds to stimuli, what it makes and where it projects, we may still not know what it does that matters to the behaving organism. By contrast, for the magnocellular vasopressin and oxytocin neurons of the hypothalamus, we can know much of what they do even before we know how they do it. All of these neurons project to the posterior pituitary gland, and what they secrete from there is measurable in the blood, and has measurable consequences for important physiological functions.
Neuroendocrinology began as the study of the secretion of peptides from neurons into the blood, and evolved to be also the study of secretion of peptides within the brain. In both aspects, electrophysiological recordings from single neurons in vivo have been fundamental in defining the physiological significance of mechanisms that have been established by cellular and molecular studies in vitro (Figure 1.1).
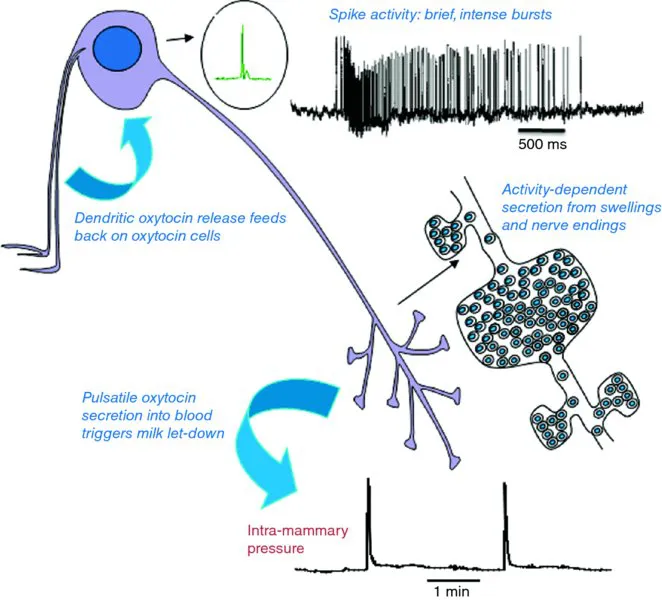
Figure 1.1The milk-ejection reflex. The reflex was uncovered by electrophysiological studies in vivo. In response to suckling, oxytocin cells discharge intermittently in brief synchronized bursts that evoke secretion of pulses of oxytocin, which induce abrupt episodes of milk ejection. Dendritically released oxytocin facilitates the bursting.
1.2 Opening the window on the brain
Single-cell recording has strengths and weaknesses—and in vivo studies have particular problems as well as certain unique advantages. Because in vivo electrophysiology requires an extensive and prolonged investment in skills and expertise, it is wise to understand the nature of the investment that is involved, as well as the potential returns and the likely limitations.
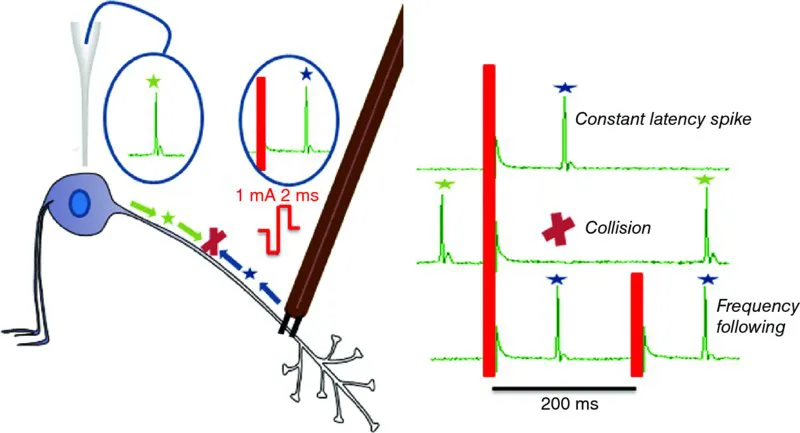
The window on the brain afforded by the magnocellular system was opened by Wakerley and Lincoln (1973), when they used a technique to enable the magnocellular neurosecretory neurons to be rigorously interrogated electrophysiologically. There had been earlier attempts to study these neurons electrophysiologically, but despite careful stereotactical control, these recordings came from a mixture of neuroendocrine and non-neuroendocrine cells. This heterogeneity subverted interpretation: the breakthrough came from the ability to identify individual neurons as neuroendocrine neurons immediately and unequivocally. That breakthrough came with the introduction of antidromic identification (Figure 1.2).
Figure 1.2Antidromic identification. A stimulating electrode that is placed on the axon of any neuron may be used to trigger a spike that is propagated both orthodromically (green stars), toward the axonal endings, and antidromically (blue stars), toward the cell body. A recording electrode at the cell body will record the antidromic spike at a fixed latency following the stimulus—a latency that reflects the conduction velocity and the axonal length. In general, a stimulus pulse might evoke a spike that is generated by monosynaptic excitation, which would also arise at a nearly constant latency. Antidromic spikes can be distinguished from such orthodromic spikes by two additional tests: frequency following and the collision test. Antidromic spikes will be generated (i.e., will follow) each of a short train of stimulus pulses presented at a high frequency (50–100 Hz); these spikes will maintain a near-constant latency (there is a slight prolongation of latency with each successive pulse). A longer train of stimuli will result in fractionation of the antidromic spike—as the soma becomes refractory to antidromic stimulation, antidromic invasion is progressively delayed and may intermittently fail, while the smaller initial segment spike, which is normally hidden within the soma spike, will be preserved and become visible as a notch on the rising phase of the antidromic spike. However, antidromic spikes will not invade either the initial segment or the soma when they are extinguished by collision with a spontaneous, orthodromically propagated spike. This collision (red X) occurs when an antidromic stimulus pulse immediately follows a spontaneously generated spike—the descending spontaneous orthodromic spike meets the ascending antidromic spike along the axon, and both are extinguished by this collision.
Because magnocellular neurons project to the pituitary, a stimulating electrode placed on the neural stalk can be used to trigger action potentials (spikes) in their axons, and these spikes can be detected at the soma, after antidromic (i.e., backward) propagation, as spikes that follow each stimulus at a constant latency. The rat supraoptic nucleus contains only two types of neuron—magnocellular oxytocin neurons and magnocellular vasopressin neurons, so every neuron recorded from this region that can be antidromically stimulated can be identified as one of these two types. Once Wakerley and Lincoln began to apply this approach, they saw that these two cell types could be distinguished by other tests. These tests were refined by many others, leading to an extensive battery of tests that now allows us to talk of identified oxytocin neurons and identified vasopressin neurons.
From the ability to identify oxytocin and vasopressin neurons in vivo came the cornerstones of our understanding of the physiology of these systems. Wakerley and Lincoln (1973) defined the milk-ejection reflex, as it is known in all good text books (Figure 1.1). From their studies came the realization that, in response to suckling, oxytocin is not released continuously, but intermittently—in pulses that result from the brief intense synchronized bursts of action potentials generated by the oxytocin neurons (Figure 1.3). From Francois Moos and her colleagues (see Richard et al., 1997), we learned that this reflex depends upon central release of oxytocin, and could be facilitated by injecting tiny amounts of oxytocin introcerebroventricularly (i.c.v.)—the first (and still the best) example of an indispensable physiological role of central peptide release. From Summerlee and Lincoln (1981), we learned that the milk-ejection reflex in conscious rats is the same as in anaesthetized rats—and that during parturition, oxytocin is again released in intermittent pulses as a result of similar synchronized bursting (Summerlee, 1981). From Jon Wakerley, Richard Dyball, and Dominique Poulain came the recognition that in response to osmotic challenge, vasopressin neurons fire not continuously, but phasically (Figure 1.4). This phasic firing is asynchronous among the vasopressin neurons, so that it leads to continuously maintained secretion—its significance is not in producing a phasic pattern of secretion, but in optimizing the efficiency of stimulus–secretion coupling at the nerve terminals. From Dyball and others, we learnt that oxytocin neurons are just as responsive as vasopressin neurons to osmotic pressure increases, leading to recognition of oxytocin's role (in many species) in regulating natriuresis. These pioneers laid the cornerst...
Table of contents
Cover
Titlepage
Copyright
List of Contributors
Series Preface
Preface
About the Companion Website
SECTION 1A Magnocellular Neuroendocrine Neurons: Properties and Control of Vasopressin and Oxytocin Neurons
SECTION 1B Magnocellular Neuroendocrine Neurons: Synaptic Plasticity and the Autoregulation of Vasopressin and Oxytocin Release
SECTION 2 Gonadotropin-Releasing Hormone Neurons of the Parvocellular Neurosecretory System
Glossary
Index
End User License Agreement
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Neurophysiology of Neuroendocrine Neurons by William E. Armstrong,Jeffrey G. Tasker in PDF and/or ePUB format, as well as other popular books in Scienze biologiche & Neuroscienza. We have over 1.5 million books available in our catalogue for you to explore.