Introduction to Pain and its relation to Nervous System Disorders provides an accessible overview of the latest developments in the science underpinning pain research, including, but not limited to, the physiological, pathological and psychological aspects. This unique book fills a gap in current literature by focussing on the intricate relationship between pain and human nervous system disorders such as Autism, Alzheimer Disease, Parkinson's Disease, Depression and Multiple Sclerosis. This fully illustrated, colour handbook will help non-experts, including advanced undergraduate and new postgraduate students, become familiar with the current, wide-ranging areas of research that cover every aspect of the field from chronic and inflammatory pain to neuropathic pain and biopsychosocial models of pain, functional imaging and genetics.
Contributions from leading experts in neuroscience and psychiatry provide both factual information and critical points of view on their approach and the theoretical framework behind their choices. An appreciation of the strengths and weaknesses of brain imaging technology applied to pain research in humans provides the tools required to understand current cutting edge literature on the topic. Chapters covering placebo effects in analgesia and the psychology of pain give a thorough overview of cognitive, psychological and social influences on pain perception. Sections exploring pain in the lifecycle and in relation to nervous system disorders take particular relevance from a clinical point of view. Furthermore, an intellectually stimulating chapter analysing the co-morbidity of pain and depression provides a philosophical angle rarely presented in related handbooks. The references to external research databases and relevant websites aim to prompt readers to become critical and independent thinkers, and motivate them to carry out further reading on these topics.
Introduction to Pain and its relation to Nervous System Disorders is essential reading for advanced undergraduate and postgraduate students in neuroscience, medical and biomedical sciences, as well as for clinical and medical healthcare professionals involved in pain management.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Primary afferent axons belonging to the somatosensory system can respond to a range of mechanical, thermal and chemical stimuli. Many of these afferents are activated by stimuli that damage (or threaten to damage) tissues, and these are known as nociceptors. Primary afferents that innervate the limbs and the trunk enter the spinal cord through the dorsal roots and form excitatory (glutamatergic) synapses with neurons in the dorsal horn. The dorsal horn contains a large number of neurons, the great majority of which have axons that arborise locally and remain in the spinal cord; these are known as interneurons and are involved in the local processing of sensory information. In addition, the dorsal horn contains projection cells – that is, neurons with axons that enter the white matter and travel rostrally to the brain. The axons of these cells are grouped into a number of different ascending tracts. The final neuronal component consists of descending axons, which originate from cells in the brain (particularly the brainstem) and terminate diffusely within the dorsal horn.
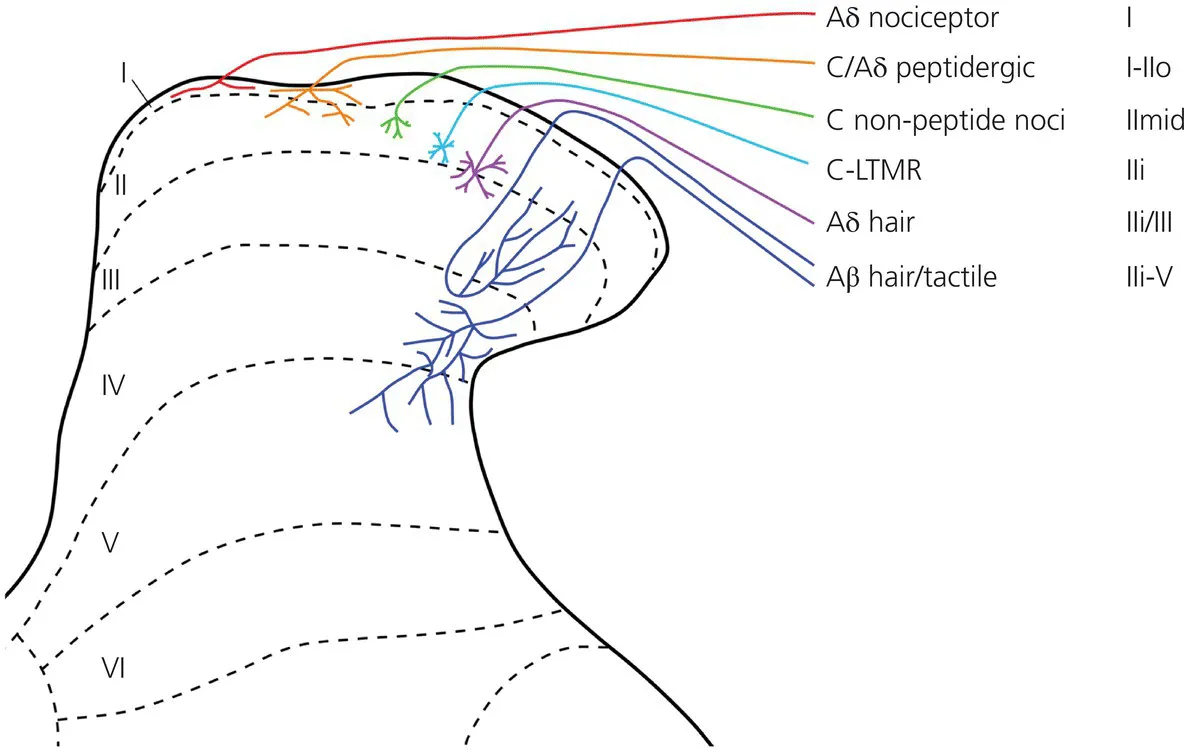
Rexed (1952) divided the grey matter of the dorsal horn into six parallel laminae (numbered from dorsal to ventral), and this scheme is widely used, for example to describe the arborisation of primary afferents and the distribution of different populations of spinal neurons (Figure 1.1). The dorsal horn is somatotopically organised, the body being mapped in a bidimensional pattern onto the rostrocaudal and mediolateral axes. The other, dorsoventral dimension is arranged in a modality-specific pattern, as will be described later.
Figure 1.1 Rexed’s laminae and primary afferent inputs. The diagram shows the main areas targeted by the central arbors of different types of primary afferent, apart from proprioceptors. These have been superimposed on Rexed’s laminar scheme, as applied to the rat dorsal horn. Note that Woodbury and Koerber (2003) have described another class of myelinated nociceptor, with axonal arbors that extend throughout laminae I–V, and this has not been included.
Source: Todd, 2010. Reproduced with permission of Nature Publishing Group.
Fifty years ago, Melzack and Wall (1965) proposed that neurons within the superficial dorsal horn could ‘gate’ the inputs from nociceptors and thus modify the perception of pain. This theory attracted a great deal of interest, and there have been numerous attempts to unravel the neuronal circuitry that underlies pain processing in the spinal cord. It turns out that this circuitry is highly complex, and we still have only a limited understanding of it. Nonetheless, it is clear that this region is very important for modulating pain in both normal and pathological states. The superficial dorsal horn is also likely to provide important targets for new drugs designed to treat pain, as it contains a wealth of receptors and signalling molecules. This chapter will summarise some of the available information about the anatomy of spinal somatosensory pathways, placing particular emphasis on the organisation of neuronal populations and their synaptic connections.
Primary afferent input to the dorsal horn
Primary afferents have their cell bodies in the dorsal root ganglia and an axon that bifurcates, sending one branch to a peripheral target and the other to the spinal cord. They can be classified according to a series of criteria. On the basis of their peripheral targets, they can be divided into afferents that innervate the skin, the muscles, the viscera and so on. Secondly, they can be characterised in terms of the strength and type of the adequate stimulus (e.g., hot/cold nociceptors, low-threshold mechanoreceptors). They also vary in axonal diameter – which is related to conduction velocity – and in whether or not they are myelinated: very fine afferents are unmyelinated and are known as C fibres, while the remainder can be divided into large-diameter (Aβ) and small-diameter (Aδ) myelinated fibres. Finally, there are various neurochemical markers, for example neuropeptides, that can be used to distinguish functional populations. These parameters are interrelated; for example, the majority of C and Aδ fibres are nociceptors or thermoreceptors, whereas most Aβs are low-threshold mechanoreceptors (LTMRs) that respond to touch or hair movement. However, this is not an absolute distinction and there are LTMRs among both the Aδ and the C fibre classes, while some nociceptors conduct action potentials in the Aβ range.
Although the focus of this chapter is on pain, the LTMRs are relevant because in many pathological pain states touch can elicit pain (tactile allodynia), and at least a part of this effect is mediated by the low-threshold afferents (Campbell et al., 1988).
The laminar distribution pattern of the main classes of primary afferent is shown in Figure 1.1.
Termination of nociceptors within the dorsal horn
Myelinated nociceptors (mostly Aδ fibres) convey ‘fast’ pain, whereas the nociceptive C fibres underlie ‘slow’ pain. The central projections of Aδ nociceptors have been demonstrated through intra-axonal labelling techniques. Many of these fibres terminate in a compact distribution in lamina I and in the outermost part of lamina II, while some arborise diffusely throughout laminae I–V (Light and Perl, 1979, Woodbury and Koerber, 2003). The central projections of myelinated afferents have also been studied through bulk-labelling after the injection of cholera toxin B subunit (CTb) into peripheral nerves. However, although this technique reveals projections to lamina I, some Aδ afferents (e.g., those terminating throughout laminae I–V) are not labelled.
Because of the small size of C fibres, there have been few studies of the central projections of individual afferents of this type (Sugiura, Lee and Perl, 1986). Conveniently, there are neurochemical markers that can be used to recognise different functional populations. C nociceptors can be divided into those that contain neuropeptides – the peptidergic group – and those that do not – the non-peptidergic group. All of the peptidergic afferents appear to contain calcitonin gene-related peptide (CGRP), which is only found in primary afferents in the dorsal horn. In addition, they can express a variety of other peptides, including substance P. Such afferents, which innervate most tissues of the body (including the skin), can be identified in anatomical experiments through immunocytochemistry for CGRP, although this approach does not distinguish between C fibres and peptidergic Aδ nociceptors (Lawson, Crepps and Perl, 1997). Most peptidergic afferents project to lamina I and the outer part of lamina II (IIo), but some send branches to deeper laminae (III–V) (Todd, 2010).
The non-peptidergic C nociceptors have been identified by their ability to bind the plant lectin IB4, although this property is not restricted to the non-peptidergic C afferents. More recently they have been shown to express mas-related G protein-coupled receptor D (MRGD) (Zylka, Rice and Anderson, 2005). These afferents innervate the skin and terminate more superficially than cutaneous peptidergic fibres. Their central arbors occupy a narrow band in the middle part of lamina II.
Central terminations of low threshold mechanoreceptors
Studies of intra-axonally injected Aβ afferents have shown that these terminate in the deep laminae, with specific patterns for the various types of tactile and hair follicle afferent. Low-threshold mechanoreceptive Aδ fibres correspond to down-hair (D-hair) afferents, and these have a more restricted projection, to the inner half of lamina II (IIi) and the dorsal part of lamina III. Recent studies have identified neurochemical/genetic markers for these afferents in the mouse, and these markers have confirmed the distribution patterns that were previously reported from intra-axonal injection studies (Abraira and Ginty, 2013). Some C fibres respond to tactile stimuli (C-LTMRs), and these have been shown to terminate in lamina IIi (Seal et al., 2009).
Vesicular glutamate transporters and primary afferents
All primary afferents use glutamate as their principal fast transmitter, and this is concentrated into synaptic vesicles by a family of three vesicular glutamate transporters (VGLUT1–3). These are differentially distributed among the primary afferents. All myelinated LTMRs express VGLUT1 and form the main source of VGLUT1 in the dorsal horn, although some originates from descending corticospinal axons (Todd et al., 2003). In contrast, Aδ nociceptors possess VGLUT2.
Both peptidergic and non-peptidergic C nociceptors express VGLUT2 (Brumovsky, Watanabe and Hokfelt, 2007), but the level of protein detected with immunocytochemistry in their central terminals is generally very low. The C-LTMRs are unique, in that they express VGLUT3 (Seal et al., 2009).
Receptors expressed by primary afferents
Primary afferents can express a wide variety of ligand-gated ionotropic, metabotropic and tyrosine kinase receptors (Todd and Koerber, 2012). For example, peptidergic and non-peptidergic C nociceptors have different growth factor receptors: trkA and RET, respectively. Glutamate receptors of the NMDA and AMPA type are widely expressed, as are GABAA and GABAB receptors, while non-peptidergic nociceptors possess purinergic P2X3 receptors. There are several receptors for neuropeptides, including μ, δ and κ opioid recep...
Table of contents
Cover
Title Page
Table of Contents
List of contributors
Foreword
Acknowledgements
Notes on authors
Introduction
SECTION I: Neurobiology of pain
SECTION II: Pain in the brain
SECTION III: Pain in the lifecycle and in nervous system disorders
APPENDIX: Interviews with chronic pain patients
Index
End User License Agreement
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access An Introduction to Pain and its relation to Nervous System Disorders by Anna A. Battaglia in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Neuroscience. We have over 1.5 million books available in our catalogue for you to explore.