This comprehensive textbook is the key resource for postgraduate trainees or residents in haematology. Now in its seventh edition, the book continues to provide everything the reader needs for examination preparation or clinical practice.
Postgraduate Haematology discusses up-to-date knowledge of the pathogenesis, clinical and laboratory features, management and treatment of a wide range of blood and bone marrow disorders in a concise and user friendly style. It presents essential information for everyday use and teaching, as well more detailed scientific background for more in-depth reading, accompanied by thoughtful referencing.
The clearly illustrated full-colour figures and charts demonstrate key facts, and are supplemented by numerous high quality photomicrographs of blood cells and tissues. Over 51 chapters from international authors, including the WHO Classification of Haematopoietic and Lymphoid Tissues, Postgraduate Haematology provides an expert review of malignant and non-malignant haematology.
New sections reflect advances in the specialty, e.g. knowledge gained from new generation sequencing, latest anticoagulant drugs, diagnostic laboratory tools, and treatment strategies
Superb four-color illustrations and photomicrographs of blood cells and tissues throughout
Includes algorithms to aid with decision-making for treatment
Companion website featuring figures and tables from the book
Arm yourself with the textbook of choice for trainees and practitioners in haematology.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Erasmus Stem Cell Institute, Erasmus Medical Centre, Rotterdam, Netherlands and University of Edinburgh, Centre for Inflammation Research, UK
Introduction
Haemopoietic stem cells (HSCs) are the foundation of the adult blood system and sustain the lifelong production of all blood lineages. These rare cells are generally defined by their ability to self-renew through a process of asymmetric cell division, the outcome of which is an HSC and a differentiating cell. In health, HSCs provide homeostatic maintenance of the system through their ability to differentiate and generate the hundreds of millions of erythrocytes and leucocytes needed each day. In trauma and physiological stress, HSCs ensure the replacement of the lost or damaged blood cells. The tight regulation of HSC self-renewal ensures the appropriate balance of blood cell production. Perturbation of this regulation and unchecked growth of HSCs and/or immature blood cells results in leukaemia. Over the last 50 years, great success has been achieved with bone marrow transplantation as a stem cell regenerative therapy. However, insufficient numbers of HSCs are still a major constraint in clinical applications. As the pivotal cells in this essential tissue, HSCs are the focus of intense research to: (1) further our understanding of their normal behaviour and the basis of their dysfunction in haemopoietic disease and leukaemia and (2) provide insights for new strategies for improved and patient-specific stem cell therapies. This chapter provides current and historical information on the organization of the adult haemopoietic cell differentiation hierarchy, the ontogeny of HSCs, the stromal microenvironment supporting these cells, and the molecular mechanisms involved in the regulation of HSCs.
Hierarchical organization and lineage relationships in the adult haemopoietic system
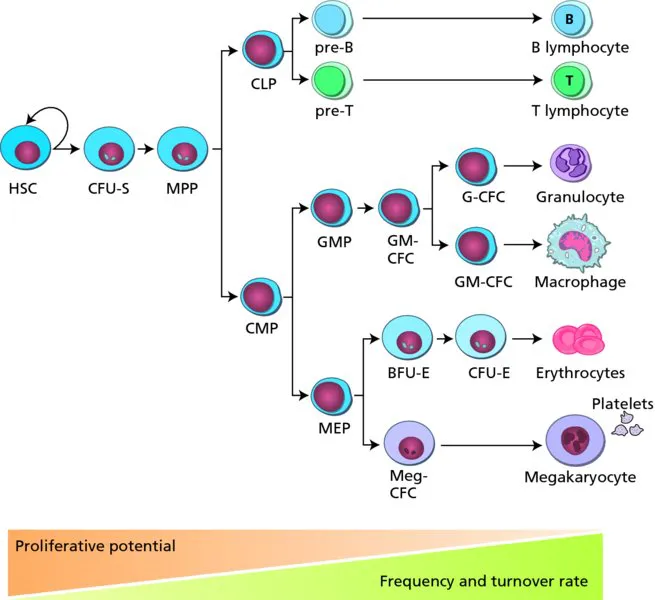
The haemopoietic system is the best-characterized cell lineage differentiation hierarchy and, as such, has set the paradigm for the growth and differentiation of tissue-specific stem cells. HSCs are defined by their high proliferative potential, ability to self-renew and potential to give rise to all haemopoietic lineages. HSCs produce immature progenitors that gradually and progressively, through a series of proliferation and differentiation events, become restricted in lineage differentiation potential. Such restricted progenitors produce the terminally differentiated functional blood cells.
The lineage relationships of the variety of cells within the adult haemopoietic hierarchy (Figure 1.1) are based on results of in vivo transplantation assays in irradiated/myeloablated recipient mice and many in vitro differentiation assays that became available following the identification of haemopoietic growth factors. These assays facilitated measurement of the maturational progression of stem cells and progenitors, at or near the branch points of lineage commitment. Clonal analyses, in the form of colony-forming unit (CFU) assays or single cell transplantation assays, were developed to define the lineage differentiation potential of the stem cell or progenitor, and to quantitate the number/frequency of such cells in the population as a whole. In general, the rarer a progenitor is and the greater its lineage differentiation potential, the closer it is in the hierarchy to the HSC. In vitro clonogenic assays measure the most immature progenitor CFU-GEMM/Mix (granulocyte, erythroid, macrophage, megakaryocyte), bipotent progenitors CFU-GM (granulocyte, macrophage) and restricted progenitors CFU-M (macrophage), CFU-G (granulocyte), CFU-E (erythroid) and BFU-E (burst-forming unit-erythroid). While such in vitro clonogenic assays measure myeloid and erythroid potential, lymphoid potential is revealed only in fetal thymic organ cultures and stromal cell cocultures in which the appropriate microenvironment and growth factors are present. Long-term culture assays (6–8 week duration), such as the cobblestone-area-forming cell (CAFC) and the long-term culture-initiating cell (LTC-IC) assays, reveal the most immature of haemopoietic progenitors. Currently, the major hurdle in studies and clinical applications of HSCs is the fact that HSCs cannot be expanded and are poorly maintained in culture. The only way to detect a bona fide HSC is in vivo.
Figure 1.1 The adult haemopoietic hierarchy. Haemopoietic stem cells are at the foundation of the hierarchy. Through a series of progressive proliferation and differentiation steps the mature blood cell lineages are produced. Haemopoietic stem cells have the greatest proliferative and multilineage differentiation potential, while the mature blood cells are not proliferative and are lineage restricted. While large numbers of mature cells are found in the blood and turn over rapidly, the bone marrow contains long-lived quiescent haemopoietic stem cells at a very low frequency.
In vivo, the heterogeneity of the bone marrow population of immature progenitors and HSCs is reflected in the time periods at which different clones contribute to haemopoiesis. Short-term in vivo repopulating haemopoietic progenitor cells such as CFU-S (spleen) give rise to macroscopic erythro-myeloid colonies on the spleen within 14 days of injection. Bona fide HSCs give rise to the long-term high-level engraftment of all haemopoietic lineages. Serial transplantations reveal the ability of the long-term repopulating HSCs to self-renew. The clonal nature of engraftment and the multilineage potential of HSCs has been demonstrated through radiation, retroviral and bar-code marking of bone marrow cells. Such studies suggest that, at steady state, several HSC clones contribute to the haemopoietic system at any one time. Further analyses of bone marrow HSCs show that this compartment consists of a limited number of distinct HSC subsets, each with predictable behaviours, as described by their repopulation kinetics in myeloablated adult recipients. In general, the bone marrow haemopoietic cell compartment, as measured by in vitro clonogenic assays and in vivo transplantation assays, shows a progression along the adult differentiation hierarchy from HSCs to progenitors and fully functional blood cells with decreased multipotency and proliferative potential.
The use of flow cytometry to enrich for HSCs and the various progenitors in adult bone marrow has been instrumental in refining precursor–progeny relationships in the adult haemopoietic hierarchy. HSCs are characteristically small ‘blast’ cells, with a relatively low forward and side light scatter and low metabolic activity. Both mouse and human HSCs are negative for expression of mature haemopoietic lineage cell-surface markers, such as those found on B lymphoid cells (CD19, B220), T lymphoid cells (CD4, CD8, CD3), macrophages (CD15, Mac-1) and granulocytes (Gr-1). Positive selection for mouse HSCs relies on expression of Sca-1, c-kit, endoglin and CD150 markers and for human HSCs on expression of CD34, c-kit, IL-6R, Thy-1 and CD45RA markers. Similarly, cell types at lineage branch points have been identified, including the CMP (common myeloid progenitor), CLP (common lymphoid progenitor) and GMP (granulocyte macrophage progenitor). Recently, using the Flt3 receptor tyrosine kinase surface marker along with many other well-studied markers, the LMPP (lymphoid primed multipotent progenitor) has been identified within the lineage negative, Sca-1 positive, c-kit positive (LSK) enriched fraction of HSCs. These cells have granulocyte/macrophage, B lymphoid and T lymphoid potential, but little or no megakaryocyte/erythroid potential. This suggests that the first lineage differentiation event is not a strict separation into common lymphoid and myeloid pathways. While these cell-surface marker changes and functional restriction events are represented by discrete cells in the working model of the haemopoietic hierarchy as depicted in textbooks and Figure 1.1, it is most likely that there is a continuum of cells between these landmarks and/or alternative differentiation paths. The currently identified progenitor cells in the hierarchy represent the cells present at stable and detectable frequencies and for which we currently have markers and functional assays. As more cell-surface markers are identified and the sensitivity of detection is increased, additional intermediate cell subsets are likely to be identified. Together with single cell transcriptomic approaches, it may be possible to predict the molecular events needed for the HSC state and the differentiation of the entire haemopoietic system.
Sites of adult haemopoiesis
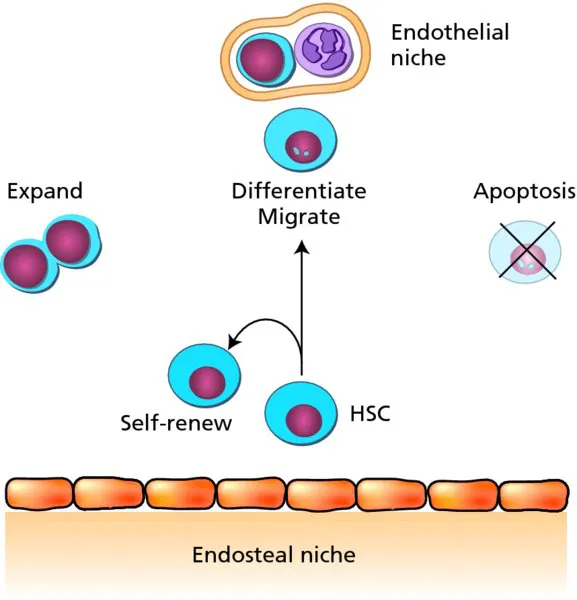
Bone marrow, spleen, thymus and lymph nodes are the haemopoietic sites in the adult, and each tissue plays a special role in supporting the growth and differentiation of particular haemopoietic cell lineages and subsets. Equally important is the blood itself, which is a mobile haemopoietic tissue, with mature blood cells travelling through the circulation to function in all parts of the body. Not only do the terminally differentiated cells, such as erythrocytes and lymphocytes, move by means of the circulation, but HSCs (at low frequency) also migrate through the circulation from the bone marrow to other haemopoietic tissues. HSCs are mostly concentrated in the bone marrow and are found in the endosteal and vascular niches (Figure 1.2). HSCs can be induced to circulate by administration of granulocyte colony-stimulating factor (G-CSF). Recent improvements in confocal microscopy have allowed the visualization of the migration of circulating HSCs to the bone marrow endosteal niche by time-lapse imaging in the mouse.
Figure 1.2 The bone marrow haemopoietic niches. Haemopoietic stem cells are found in the endosteal and endothelial niches of the bone marrow. These niches support the maintenance, self-renewal, expansion, differentiation, migration and survival of haemopoietic stem cells through local growth factor production and cell–cell interactions.
The estimated frequency of HSCs is 1 per 104–105 mouse bone marrow cells and 1 per 20 × 106 human bone marrow cells. HSCs are also found in the mouse spleen at approximately a 10-fold lower frequency and in the circulating blood at a 100-fold lower frequency. The capacity for HSCs to migrate and also be retained in bone marrow supportive niches is of relevance to clinical transplantation therapies. HSCs injected intravenously in such therapies must find their way to the bone marrow for survival and effective haemopoietic engraftment. For example, stromal-derived factor (SDF)-1 and its receptor CXCR4 (expressed on HSCs) are implicated in the movement of HSCs and the retention of HSCs in the bone marrow. Indeed, HSC mobilization can be induced through AMD3100, an antagonist of SDF-1, and by the administration of G-CSF. Mobilization strategies with G-CSF are used routinely to stimulate bone marrow HSCs to enter the circulation, allowing ease of collection in the blood rather than through bone marrow biopsy.
Development of HSCs
Waves of haemopoietic generation in embryonic development
Until the mid-1960s it was thought that blood cells were intrinsically generated in tissues such as the liver, spleen, bone marrow and thymus. Survival studies in which cells from un-irradiated tissues were injected into lethally irr...
Table of contents
Cover
Companion website
Title page
Copyright
Contributor list
Preface to the seventh edition
Preface to the first edition
Chapter 1 Stem cells and haemopoiesis
Chapter 2 Erythropoiesis
Chapter 3 Iron metabolism, iron deficiency and disorders of haem synthesis
Chapter 4 Iron overload
Chapter 5 Megaloblastic anaemia
Chapter 6 Haemoglobin and the inherited disorders of globin synthesis
Chapter 7 Sickle cell disease
Chapter 8 Hereditary disorders of the red cell membrane and disorders of red cell metabolism
Chapter 23 Supportive care in the management of leukaemia
Chapter 24 Chronic myeloid leukaemia
Chapter 25 The myelodysplastic syndromes
Chapter 26 Myeloproliferative neoplasms
Chapter 27 Chronic lymphocytic leukaemia and other chronic B-cell disorders
Chapter 28 T-cell lymphoproliferative disorders
Chapter 29 Multiple myeloma
Chapter 30 Amyloidosis
Chapter 31 The classification of lymphomas: updating the WHO classification
Chapter 32 Hodgkin lymphoma
Chapter 33 Non-Hodgkin lymphoma: low grade
Chapter 34 Non-Hodgkin lymphoma: high grade
Chapter 35 Stem cell transplantation
Chapter 36 Normal haemostasis
Chapter 37 The vascular function of platelets
Chapter 38 Haemophilia and Von Willebrand Disease
Chapter 39 Rare inherited coagulation disorders
Chapter 40 Acquired coagulation disorders
Chapter 41 Congenital platelet disorders
Chapter 42 Primary immune thrombocytopenia
Chapter 43 Thrombotic thrombocytopenic purpura and haemolytic–uraemic syndrome (congenital and acquired)
Chapter 44 Heritable thrombophilia
Chapter 45 Acquired venous thrombosis
Chapter 46 Antithrombotic agents
Chapter 47 Management of venous thromboembolism
Chapter 48 Haematological aspects of systemic disease
Chapter 49 Haematological aspects of tropical diseases
Chapter 50 Neonatal haematology
Chapter 51 WHO Classification: Tumours of the Haematopoietic and Lymphoid Tissues (2008)
Index
EULA
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Postgraduate Haematology by Victor Hoffbrand, Douglas R. Higgs, David M. Keeling, Atul B. Mehta, A. Victor Hoffbrand,Douglas R. Higgs,David M. Keeling,Atul B. Mehta, A. Victor Hoffbrand, Douglas R. Higgs, David M. Keeling, Atul B. Mehta in PDF and/or ePUB format, as well as other popular books in Medicine & Hematology. We have over 1.5 million books available in our catalogue for you to explore.