The Ichneumonoidea is a vast and important superfamily of parasitic wasps, with some 60,000 described species and estimated numbers far higher, especially for small-bodied tropical taxa. The superfamily comprises two cosmopolitan families - Braconidae and Ichneumonidae - that have largely attracted separate groups of researchers, and this, to a considerable extent, has meant that understanding of their adaptive features has often been considered in isolation. This book considers both families, highlighting similarities and differences in their
adaptations.
The classification of the whole of the Ichneumonoidea, along with most other insect orders, has been plagued by typology whereby undue importance has been attributed to particular characters in defining groups. Typology is a common disease of traditional taxonomy such that, until recently, quite a lot of taxa have been associated with the wrong higher clades. The sheer size of the group, and until the last 30 or so years, lack of accessible identification materials, has been a further impediment to research on all but a handful of 'lab rat' species usually cultured initially because of their potential in biological control.
New evidence, largely in the form of molecular data, have shown that many morphological, behavioural, physiological and anatomical characters associated with basic life history features, specifically whether wasps are ecto- or endoparasitic, or idiobiont or koinobiont, can be grossly misleading in terms of the phylogeny they suggest. This book shows how, with better supported phylogenetic hypotheses entomologists can understand far more about the ways natural selection is acting upon them.
This new book also focuses on this superfamily with which the author has great familiarity and provides a detailed coverage of each subfamily, emphasising anatomy, taxonomy and systematics, biology, as well as pointing out the importance and research potential of each group. Fossil taxa are included and it also has sections on
biogeography, global species richness, culturing and rearing and preparing specimens for taxonomic study. The book highlights areas where research might be particularly rewarding and suggests systems/groups that need investigation. The author provides a large compendium of references to original research on each group. This book is an essential workmate for all postgraduates and researchers working on ichneumonoid or other parasitic wasps worldwide. It will stand as a reference book for a good number of years, and while rapid advances in various fields such as genomics and host physiological interactions will lead to new information, as an overall synthesis of the current state it will stay relevant for a long time.

eBook - ePub
The Braconid and Ichneumonid Parasitoid Wasps
Biology, Systematics, Evolution and Ecology
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
The Braconid and Ichneumonid Parasitoid Wasps
Biology, Systematics, Evolution and Ecology
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 1
Introduction
Although most people are blissfully unaware of them, the ichneumonoid wasps are one of the most diverse groups of insects, and in terms of their ecological role they are probably of enormous importance. No-one really has a good idea about how diverse they are and estimates vary widely. The total number of valid species described to date, 18,000 braconids and 23,000 ichneumonids1, is certainly a great underestimate, but by how much is still anyone's guess. Many works cite estimates of 40,000 and 60,000, based upon expert opinion (Townes 1969, Gauld & Bolton 1988). Similar values have also been obtained by various objective estimation measures, but it seems likely that these too are underestimates, and narrowing the numbers down is not going to be easy for the reasons explained in Chapter 15.
Unfortunately, neither family has attracted a lot of attention from amateur entomologists, which seems to be a prerequisite for a good knowledge of a group's taxonomy, distribution and biology. This may be partly because many of the species are rather small and often dull coloured, although this does not seem to have deterred generations of amateur coleopterists. Probably the most important factor has been the dearth, until fairly recently, of reliable and accessible identification guides to the major groups (subfamilies), confounded by the fact that the subfamily-level classification is only now becoming fairly stable, largely as a result of much new molecular work. Problems have been compounded because numerous names were mis-applied by early workers and, as these errors were slowly discovered and corrected, many groups accumulated a historical backlog of alternative names. In many fields of science, the really old literature seldom has to be cited, but in zoology, a great deal of excellent work on anatomy and biology was carried out 50 to 100 or so years ago. As this may be the only detailed work on a given group, it is still relevant today and the reader therefore has to deal with the sometimes confusing or even misleading nomenclature.
Difficulties in the correct identification of specimens, and publications dealing with incorrectly identified specimens, have also been a major stumbling blocks. To quote Perkins (1959), ‘It is perhaps, not surprising that keys to subfamilies are very imperfect, as exceptions can be found to almost all characters that have been used in defining any subfamily, even in the limited British fauna’. Partly because of overall improving taxonomic and systematic understanding, published research on both families is growing, that dealing with the braconids slightly more quickly than that for the ichneumonids (Fig. 1.1). There may be several reasons for this growth, not the least of which is that most researchers are now under great pressure to publish their findings quickly in bite-sized chunks and in high-impact journals, rather than presenting single, large tomes representing the results of many years of their work. The difference in the rate of publication between the families could well be due to the ease of identification – recognising subfamilies is generally easier for braconids and knowing what subfamily you are dealing with is the essential first step towards a proper identification.
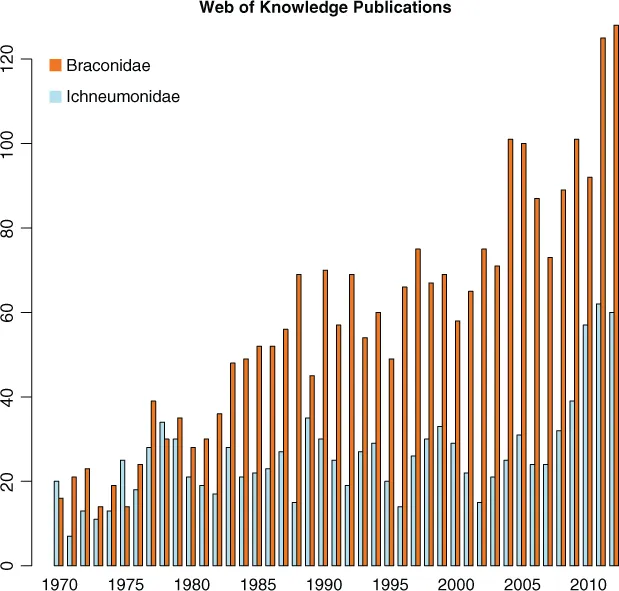
Fig. 1.1 Numbers of papers on Braconidae and Ichneumonidae published each year in Science Citation Index (SCI) journals from 1970 to 2012.
The Ichneumonidae and Braconidae are each such large groups that few people since the early 20th century have attempted to work seriously on the whole of either one of them, so it is hardly surprising that in recent years almost no-one has attempted to tackle them both. This, of course, means that the similarities and differences between them may have been less well considered than they should have been. Superficially, it might seem that these two families essentially parallel one another, they are sister groups and they broadly occupy the same range of niches—they predominantly parasitise exposed and concealed moth and beetle larvae with a few incursions into attacking fly and Hymenoptera larvae, rarer ones into other insect groups and a few other ways of life such as spider egg predation and even a few instances of true phytophagy. However, things may not be as simple as they seem, because despite some remarkable parallels, they also show strong group differences in precisely what they do and in the types of adaptations they typically employ.
It should come as no surprise therefore, that ichneumonids and braconids do not ‘behave’ in the same way in so many aspects of their biology and morphology. If they did, it seems likely that one would have driven the other to extinction or pushed them a long way in that direction. That both groups are highly speciose seems very likely to indicate that they do not compete in a precise and consistent way, although many individual species no doubt do. Hence there are various sorts of adaptations that appear to evolve frequently in one family but not or only rarely in the other. For example, numerous braconids have evolved carapace-like metasomas where the basal 3 (or sometimes 4) metasomal terga are enlarged, frequently fused, or at least more or less immovably joined and conceal all more posterior ones (see Chapter 10, section Carapacisation). Only a very few ichneumonid groups have members with carapaces and the numbers of species involved is very small. Is this associated with the difference in articulation between the second and third metasomal terga, which is one of the diagnostic features for separating the two families? Endoparasitoid larvae belonging to several different braconid lineages have apparently independently evolved an everted rectum forming a structure called an anal vesicle (see Fig. 5.1) that serves a variety of physiological roles, but this adaptation, as far as is known, has only evolved in two genera within the Ichneumonidae. Similarly, very elongate mouthparts (although variously involving the glossa, malar region or maxillary palps) have evolved on numerous independent occasions within those Braconidae dwelling in relatively arid habitats (see Chapter 10, section Concealed nectar extraction apparatus), but the number of such occurrences in the Ichneumonidae is small (e.g. Rhynchobanchus: Banchinae). These modified mouthparts, collectively referred to as a concealed nectar extraction apparatus, are an adaptation to obtain nectar from plants such as Asteraceae or Dipsaciaceae, which in turn are adapted to prevent their nectar from drying up in places where water is in short supply. In this case, it may be because braconids tend to comprise a relatively larger proportion of species in such habitats, but the data are not really available to test this.
Ichneumonids collectively utilise a somewhat different spectrum of hosts than braconids. They include many more taxa that are parasitoids of other Hymenoptera, including both endo- and ectoparasitism, in addition to acting as pseudohyperparasitoids of other ichneumonoids (see Fig. 13.1; cf. Fig. 12.2), and endoparasitism including developing as true hyperparasitoids within a host, as well as some being predators within aculeate wasp and bee nests. In the Braconidae, members of two tribes within the Euphorinae are endoparasitoids on adult Hymenoptera, a few ectoparasitoids attack leaf-mining sawflies and only a few members of the Ichneutinae are endoparasitic within sawfly larvae, and Gauld (1988a) plausibly suggested that these made the transition to sawfly hosts from ancestors that were endoparasitoids of leaf-mining Lepidoptera. Further, no braconids apart from the rather special case of a few euphorines parasitising adult ichneumonoids (see Chapter 12, section Syntretini), no braconids are hyperparasitoids or even pseudohyperparasitoids. Two subfamilies within Ichneumonidae, involving several evolutionary transitions, have become associated with spiders either as egg predators or as parasitoids of juvenile and adult individuals. All of these seem to be connected by their use of silk, or volatile or non-volatile compounds associated with silk, in the host location – because of its non-solubility, silk proteins themselves seem an incredibly unlikely source of host-finding cues. Nevertheless, at least some braconids do utilise cues from host silk trails (Ha et al. 2006), but it does not seem to have become an important part of their behavioural repertoire. Perhaps partly associated with this and the places where silk-cocooned hosts occur, ichneumonids appear to have evolved vibrational sounding (a sort of echolocation) as a host location tool on multiple occasions (and lost it on many also), whereas there is no evidence for this host location mode in the Braconidae (see Chapter 10, section Antennal hammers and vibrational sounding).
Another important question that we ought to consider is why the ichneumonoids and chalcidoids have not out-competed one another in one direction or another. Some niches occupied by chalcidoids are not available to ichneumonids; for example, egg parasitism, which necessitates body sizes smaller than or at least at the very bottom range of that which ichneumonoids (e.g. Miracinae or Cheloninae–Adeliini) have thus far achieved. Ichneumonids described to date are, in general, larger bodied than braconids (see Fig. 15.6), and this may correlate with some differences in host utilisation, since only braconids can attack small insect hosts such as psocids, aphids, plant bugs and tiny beetles (Čapek 1970).
It seems to me a very great shame that many traditional areas of study, such as those on comparative embryology and detailed descriptions of natural history, have suffered a serious decline in recent years and effectively have ceased in most Western universities. For a long time they have been largely restricted to workers in parts of the world, such as the former Soviet block countries, where access to more trendy modern methods and thought were perhaps restricted. This means that many important descriptions of biology come from before World War II and sometimes before World War I. And although many of these are of high quality, they often deal with species serendipitously, as well as under unfamiliar names that have been lost in synonymy and therefore may require some detective work. However, these are the only sources of detailed biology for some groups.
A great deal of what we know about the biologies of various groups comes from efforts to use them for biological control (e.g. Wharton 1984). As a consequence, we know far more about some subfamilies than we do about others and obviously we know more about taxa that are readily easily brought into culture, which means that the host nearly always has to be easy to culture too, or at least easy to find and collect. There are a surprisingly large number of subfamilies for which we know absolutely nothing about the biology, not even the order of hosts that they attack or whether they are ecto- or endoparasitoids. Some of them include fairly common and frequently collected species.
Now that we have the powerful tool that modern phylogenetics provides, we are in desperate need of more such studies to help test hypotheses about the adaptive natures of particular character states within a comparative framework. Although it is possible in some cases to go out and obtain the necessary taxa, there is a general mismatch of people skills. Many excellent physiologists and molecular geneticists carry out their work on taxa of real or potential economic importance and a considerable amount of their research receives funding because of this. By their nature, the hosts of potential biocontrol insects are generally easy to obtain and culture, although admittedly wood-borers may pose more of a logistic issue than say grass-feeding aphids or cotton-feeding moths. The parasitoids that are therefore best investigated are those which attack these hosts with the consequence that much work has been carried out on a relatively small subset of taxa, aphidiine and microgastrine braconids and campoplegine ichneumonids being prime examples. Many of the laboratory researchers would love to obtain some other parasitoid taxa into culture to study, but this involves setting up host cultures, obtaining the parasitoids and working out rearing techniques, all probably with less funding available. Although there are numerous exceptions, many laboratory-based experimentalists often do not have the field entomological or natural history backgrounds to facilitate the finding of some of the other taxa.
The other side of the skills mismatch is that there are, at least in many ‘Western’ and East Asian countries, excellent natural historians who are good at and enjoy going into the field in search of insects and rearing them, but they often do not necessarily know what more detailed pieces of information about an insect's biology are missing. Added to this is the problem that many taxonomically interesting taxa, which might well have particularly interesting biologies and associated physiology, biochemistry, etc., are simply rare, very local in distribution or attack hosts that are very difficult to obtain or bring into culture. There are multiple examples of all of these.
One of the aspects that really needs to be revisited, as soon as sufficient independent molecular phylogenies become available, is all the hypothesised evolutionary transitions and trends that have been based on purely morphological phylogenetic estimates. It is surprising how often the networks obtained from morphological and molecular analyses are similar, which is good, but the rooting is extremely different. Such different ...
Table of contents
- Cover
- Title Page
- Copyright
- Dedication
- Preface
- Acknowledgements
- Chapter 1: Introduction
- Part I: Morphology and Biology
- Part II: Taxonomic and Systematic Treatment
- Part III: Ecology and Diversity
- Glossary
- References
- Author Index
- General Index
- Host Index
- Ichneumonoid Genus, Tribe and Subfamily Index
- Ichneumonoidea Species Index
- End User License Agreement
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Braconid and Ichneumonid Parasitoid Wasps by Donald L. J. Quicke in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Entomology. We have over 1.5 million books available in our catalogue for you to explore.