Edited by three pioneers in the field, each with longstanding experience in the biotech industry, and a skilled scientific writer, this is the first book to cover every step in the development and production of immunoglobulin Fc-fusion proteins as therapeutics for human disease: from choosing the right molecular design, to pre-clinical characterization of the purified product, through to batch optimization and quality control for large-scale cGMP production. The whole of the second part is devoted to case studies of Fc-fusion proteins that are now commercially successful products. In this section, the authors, several of whom were personally involved in clinical development of the products themselves, detail the product?s background and give insight into issues that were faced and how these issues were overcome during clinical development. This section also includes a chapter on promising new developments for the future. An invaluable resource for professionals already working on Fc-fusion proteins and an excellent and thorough introduction for physicians, researchers, and students entering the field.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Arvind Rajpal, Pavel Strop, Yik Andy Yeung, Javier Chaparro-Riggers, and Jaume Pons
1.1 Introduction to Antibodies
Antibodies, a central part of humoral immunity, have increasingly become a dominant class of biotherapeutics in clinical development and are approved for use in patients. As with any successful endeavor, the history of monoclonal antibody therapeutics benefited from the pioneering work of many, such as Paul Ehrlich who in the late nineteenth century demonstrated that serum components had the ability to protect the host by “passive vaccination” [1], the seminal invention of monoclonal antibody generation using hybridoma technology by Kohler and Milstein [2], and the advent of recombinant technologies that sought to reduce the murine content in therapeutic antibodies [3].
During the process of generation of humoral immunity, the B-cell receptor (BCR) is formed by recombination between variable (V), diversity (D), and joining (J) exons, which define the antigen recognition element. This is combined with an immunoglobulin (Ig) constant domain element (μ for IgM, δ for IgD, γ for IgG (gamma immunoglobulin), α for IgA, and
for IgE) that defines the isotype of the molecule. Sequences for these V, D, J, and constant domain genes for disparate organisms can be found through the International ImMunoGeneTics Information System® [4]. The different Ig subtypes are presented at different points during B-cell maturation. For instance, all naïve B cells express IgM and IgD, with IgM being the first secreted molecule. As the B cells mature and undergo class switching, a majority of them secrete either IgG or IgA, which are the most abundant class of Ig in plasma.
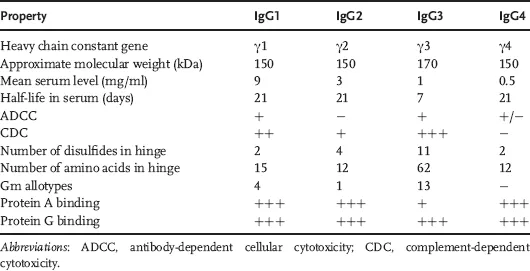
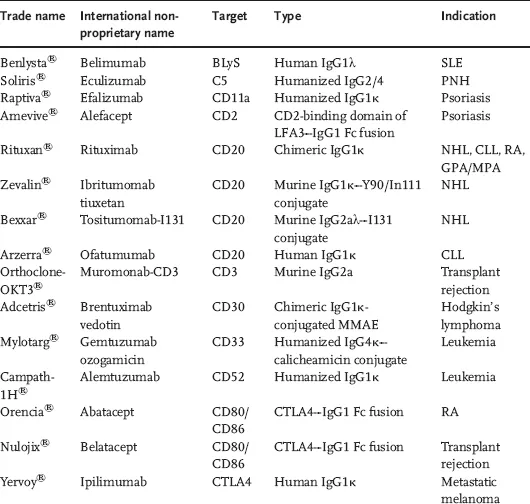
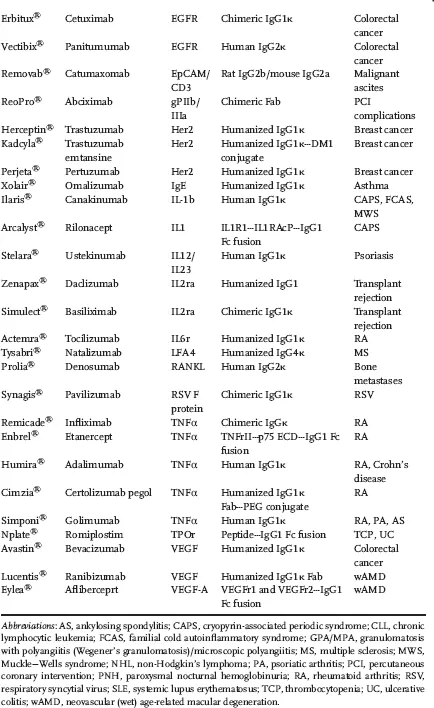
Characteristics like high neutralizing and recruitment of effector mechanisms, high affinity, and long resident half-life in plasma make the IgG isotype an ideal candidate for generation of therapeutic antibodies. Within the IgG isotype, there are four subtypes (IgG1–IgG4) with differing properties (Table 1.1). Most of the currently marketed IgGs are of the subtype IgG1 (Table 1.2).
Table 1.1 Subtype properties.
Table 1.2 Marketed antibodies and antibody derivatives by target.
The ability of antibodies to recognize their antigens with exquisite specificity and high affinity makes them an attractive class of molecules to bind extracellular targets and generate a desired pharmacological effect. Antibodies also benefit from their ability to harness an active salvage pathway, mediated by the neonatal Fc receptor (FcRn), thereby enhancing their pharmacokinetic (PK) life span and mitigating the need for frequent dosing. The antibodies and antibody derivatives approved in the United States and the European Union (Table 1.2) span a wide range of therapeutic areas, including oncology, autoimmunity, ophthalmology, and transplant rejection. They also harness disparate modes of action like blockade of ligand binding and subsequent signaling, and receptor and signal activation, which target effector functions (antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC)), and delivery of cytotoxic payload.
Antibodies are generated by the assembly of two heavy chains and two light chains to produce two antigen-binding sites and a single constant domain region (Figure 1.1, panel a). The constant domain sequence in the heavy chain designates the subtype (Table 1.1). The light chains can belong to two families (λ and κ), with most of the currently marketed antibodies belonging to the κ family.
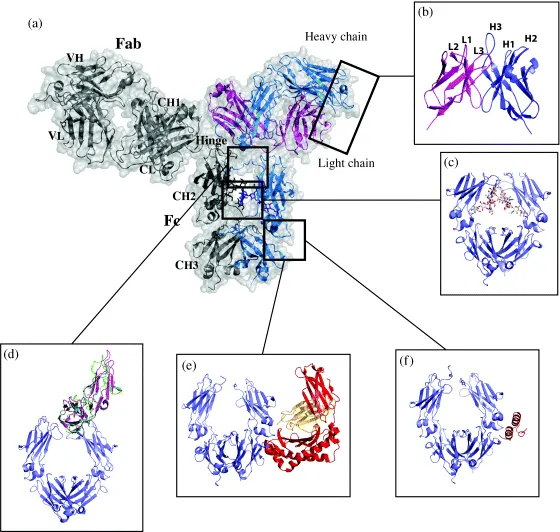
Figure 1.1 Structure and features of the IgG and its interactions. (a) The structure of a full-length IgG is shown in ribbon representation with transparent molecular surface. One heavy chain is shown in blue and one light chain in magenta. The other heavy chain and light chain are shown in gray for clarity. In this orientation, two Fab domains sit on top of the Fc domain and are connected in the middle by the hinge region. The Fab domain is composed of the heavy chain VH and CH1 domains and the light chain VL and CL domains–Protein Data Bank (PDB) [5] code 1HZH [6]. (b) Each variable domain contains three variable loops (L1–L3 on light chain and H1–H3 on heavy chain) that make up the antigen-binding site–PDB code 1HZH [6]. (c) The Fc region is composed of the dimer of CH2 and CH3 domains. The CH3 domains form a tight interaction while the CH2 domains interact through protein–protein, protein–carbohydrate, and carbohydrate–carbohydrate contacts–PDB code 1HZH [6]. (d) The hinge region is composed of a flexible region covalently tied together through disulfide bridges. Structures of the FcγRIIIa and FcγRIIa bound to the Fc are shown. The structures reveal that both receptors bind to the CH2 domain near the hinge and carbohydrates and upon their binding create an asymmetry such that the second FcγR is unable to bind. In this panel, FcγRIII is shown in green, and the FcγRII is shown in purple–PDB codes 3RY6 [7] and 1T83 [8]. (e) The crystal structure of the complex between the Fc and FcRn reveals that FcRn binds between the CH2 and CH3 domains in the Fc. FcRn chains are shown in red and orange–PDB code 1FRT [9]. (f) Interestingly, the same region also binds to bacterial Protein A commonly used for purification–PDB code 1FC2 [10].
The antigen-binding regions can be derived by proteolytic cleavage of the antibody to generate antigen-binding fragments (Fab) and the constant fragment (Fc, also known as the fragment of crystallization). The Fab comprises the variable regions (variable heavy (VH) [11] and variable light (VL)) and constant regions (CH1 and Cκ/Cλ). Within these variable regions reside loops called complementarity determining regions (CDRs) responsible for direct interaction with the antigen (Figure 1.1, panel b). Because of the significant variability in the number of amino acids in these CDRs, there are multiple numbering schemes for the variable domains [12,13] but only one widely used numbering scheme for the constant domain (including portions of the CH1, hinge, and the Fc) called the EU numbering system [14].
There are two general methods to generate antibodies in the laboratory. The first utilizes the traditional methodology employing immunization followed by recovery of functional clones either by hybridoma technology or, more recently, by recombinant cloning of variable domains from previously isolated B cells displaying and expressing the desired antigen-binding characteristics. There are several variations of these approaches. The first approach includes the immunization of transgenic animals expressing subsets of the human Ig repertoire (see review by Lonberg [15]) and isolation of rare B-cell clones from humans exposed to specific antigens of interest [16]. The second approach requires selecting from a large in vitro displayed repertoire either amplified from natural sources (i.e., human peripheral blood lymphocytes in Ref. [17]) or designed synthetically to reflect natural and/or desired properties in the binding sites of antibodies [18,19]. This approach requires the use of a genotype–phenotype linkage strategy, such as phage or yeast display, which allows for the recovery of genes for antibodies displaying appropriate binding characteristics for the antigen.
1.2 General Domain and Structure of IgG
Topologically, the IgG is composed of two heavy chains (50 kDa each) and two light chains (25 kDa each) with total molecular weight of approximately 150 kDa. Each heavy chain is composed of four domains: the variable domain (VH), CH1, CH2, and CH3. The light chain is composed of variable domain (VL) and constant domain (CL). All domains in the IgG are members of the Ig-like domain family and share a common Greek-key beta-sandwich structure with conserved intradomain disulfide bonds. The CLs contain seven strands with three in one sheet, and four in the other, while the VLs contain two more strands, resulting in two sheets of four and five strands.
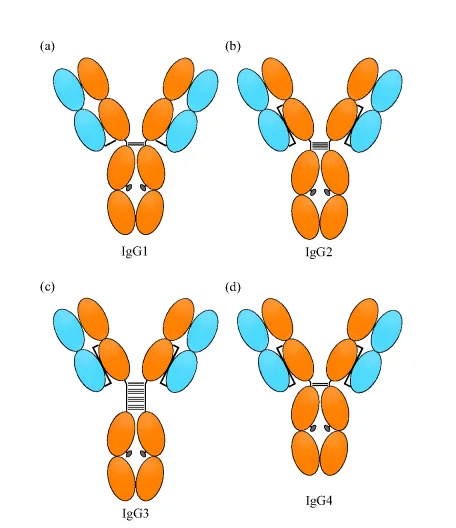
The light chain pairs up with the heavy chain VH and CH1 domains to form the Fab fragment, while the heavy chain CH2 and CH3 domains dimerize with additional heavy chain CH2 and CH3 domains to form the Fc region (Figure 1.1, panel c). The Fc domain is connected to the Fab domain via a flexible hinge region that contains several disulfide bridges that covalently link the two heavy chains together. The light chain and heavy chains are also connected by one disulfide bridge, but the connectivity differs among the IgG subclasses (Figure 1.2). The overall structure of IgG resembles a Y-shape, with the Fc region forming the base while the two Fab domains are available for binding to the antigen [6]. Studies have shown that in solution the Fab domains can adopt a variety of conformations with regard to the Fc region.
Figure 1.2 Interchain disulfide topology in human IgG subclasses. Only H–H hinge and H–L chain disulfides are shown. (a) IgG1, (b), IgG2, (c) IgG3, and (d) IgG4.
1.2.1 Structural Aspects Important for Fc Fusion(s)
1.2.1.1 Fc Protein–Protein Interactions
While the Fab region of an antibody is responsible for binding and specificity to a given target, the Fc region has many important functions outside its role as a structural scaffold. The Fc region is responsible for the long half-life of antibodies as well as for their effector functions including ADCC, CDC, and phagocytosis [20].
The long half-life of human IgGs relative to other serum proteins is a consequence of the pH-dependent interaction with the FcRn [21–23]. In the endosome, FcRn binds to the Fc region and recycles the antibody back to the plasma membrane, where the increase in pH releases the antibody back to the serum, thus rescuing it from degradation. The details of FcRn binding and its effects on antibody pharmacokinetics, including results from modulating FcRn interaction by protein engineering, are discussed in Section 1.3.3. One FcRn binds between the CH2 and CH3 domains of an Fc dimer half (Figure 1.1, panel e) [21]; therefore, up to two FcRns can bind to a single Fc.
Fc region is also responsible for binding to bacterial Protein A [10] and Protein G [24], which are commonly used for purification of Fc-containing proteins. Although Protein A binds to Fc mainly through hydrophobic interactions and Protein G through charged and polar interactions, Proteins A and G bind to a similar site on Fc domain and compete with each other (Figure 1.1, panel f). Interestingly, the binding occurs between the CH2 and CH3 domains of the Fc and largely overlaps with the FcRn binding site.
ADCC function is mediated by the interaction of the Fc region with Fcγ receptors (FcγRs). Biochemical data and structures of Fc in complex with FcγRIII and FcγRII reveal that the FcγRs bind to the combination of the Fc CH2 domain and the lower hinge region (Figure 1.1, panel d) [7,8,25]. Members of the Fcγ family have been found to bind to the same region of Fc [20,26,27] and form a 1 : 1 asy...
Table of contents
Cover
Related Titles
Title Page
Copyright
Preface
List of Contributors
Chapter 1: Introduction: Antibody Structure and Function
Part One: Methods of Production for Fc-Fusion Proteins
Part Two: Case Studies of Therapeutic Fc-Fusion Proteins
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Therapeutic Fc-Fusion Proteins by Steven M. Chamow, Thomas Ryll, Henry B. Lowman, Deborah Farson, Steven M. Chamow,Thomas Ryll,Henry B. Lowman,Deborah Farson in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biotechnology. We have over 1.5 million books available in our catalogue for you to explore.