
eBook - ePub
Fish Morphology
HiranM. Dutta
This is a test
Buch teilen
- 434 Seiten
- English
- ePUB (handyfreundlich)
- Über iOS und Android verfügbar
eBook - ePub
Fish Morphology
HiranM. Dutta
Angaben zum Buch
Buchvorschau
Inhaltsverzeichnis
Quellenangaben
Über dieses Buch
Topics covered in this volume include: transformation morphology on structures in the head of cichlid fishes; the structure and function of fish liver; atretic follicles and corpora lutea in the ovaries of fishes; effects of gill dimension on respiration; and the efects of pesticides on fish.
Häufig gestellte Fragen
Wie kann ich mein Abo kündigen?
Gehe einfach zum Kontobereich in den Einstellungen und klicke auf „Abo kündigen“ – ganz einfach. Nachdem du gekündigt hast, bleibt deine Mitgliedschaft für den verbleibenden Abozeitraum, den du bereits bezahlt hast, aktiv. Mehr Informationen hier.
(Wie) Kann ich Bücher herunterladen?
Derzeit stehen all unsere auf Mobilgeräte reagierenden ePub-Bücher zum Download über die App zur Verfügung. Die meisten unserer PDFs stehen ebenfalls zum Download bereit; wir arbeiten daran, auch die übrigen PDFs zum Download anzubieten, bei denen dies aktuell noch nicht möglich ist. Weitere Informationen hier.
Welcher Unterschied besteht bei den Preisen zwischen den Aboplänen?
Mit beiden Aboplänen erhältst du vollen Zugang zur Bibliothek und allen Funktionen von Perlego. Die einzigen Unterschiede bestehen im Preis und dem Abozeitraum: Mit dem Jahresabo sparst du auf 12 Monate gerechnet im Vergleich zum Monatsabo rund 30 %.
Was ist Perlego?
Wir sind ein Online-Abodienst für Lehrbücher, bei dem du für weniger als den Preis eines einzelnen Buches pro Monat Zugang zu einer ganzen Online-Bibliothek erhältst. Mit über 1 Million Büchern zu über 1.000 verschiedenen Themen haben wir bestimmt alles, was du brauchst! Weitere Informationen hier.
Unterstützt Perlego Text-zu-Sprache?
Achte auf das Symbol zum Vorlesen in deinem nächsten Buch, um zu sehen, ob du es dir auch anhören kannst. Bei diesem Tool wird dir Text laut vorgelesen, wobei der Text beim Vorlesen auch grafisch hervorgehoben wird. Du kannst das Vorlesen jederzeit anhalten, beschleunigen und verlangsamen. Weitere Informationen hier.
Ist Fish Morphology als Online-PDF/ePub verfügbar?
Ja, du hast Zugang zu Fish Morphology von HiranM. Dutta im PDF- und/oder ePub-Format sowie zu anderen beliebten Büchern aus Biological Sciences & Marine Biology. Aus unserem Katalog stehen dir über 1 Million Bücher zur Verfügung.
Information
1
Transformation Morphology on Structures in the Head of Cichlid Fishes
Induction and deduction are the two methods for establishing and explaining the relationship between features or for deriving one feature from another, such as form from function. Out of the two methods deduction is the strongest, while induction is used mainly in a narrative way for collecting and classifying data and, in functional morphology, for describing correlations between form and function. This description asks subsequently for a tested explanation. Deduction in functional morphology, and also in constructional and ecological morphology, aims to explain animal form and features from function, behavior, and environmental conditions. This methodological reasoning follows a reverse course. Ecological parameters are measured from which a possible or necessary behavior is derived and from this in turn a function followed by a structure. The.structures in the deductively obtained model are tested by comparing them with the actual structures (Dullemeijer and Barel, 1977).
In the following example, the structures in the facial part of cichlid fishes, we shall show how deduction can be used, where it fails or falls short, and consequently what kind of new information or models are required to be obtained in further investigations.
We shall start the procedure by presenting the course of research for the explanation of one functional component, viz. the eye, and from there proceed by adding more components to explain structures in the facial part of the head in relation to the parameters of the niche of the fish.
We shall show that structures can be explained by deduction, but also that we have often to return to induction and even have to admit that as yet the structures cannot be explained. In particular it will be demonstrated that deduction has its limitations for the following reasons. The niche parameters cannot be defined accurately, the functional capacities of the structure are insufficiently known, constraints and thus limitations occur when structures are combined, nonfunctional structures or functional for only part of the lifecycle have to be taken into account and, last but not least, some basic structural properties can only be explained historically, and thus have to be taken as given boundary conditions in the design of the model. The latter condition should force us to start with a description of structures constituting the basic plan. However, we trust that the reader will have some knowledge of the general body plan and building material of a percoid fish.
THE EYE OF CICHLID FISHES
Most cichlids have normal fish eyes, i.e., nearly hemispherical in shape with a spherical lens of which the radius (r) is about 2.55 times smaller than the radius of the retinal sphere. The lens is the only refractive element with a focal distance of 2.3 r (Fig. 1). It is situated in a myopic position to the caudal part of the retina and nearly at focal distance to the rostral part. A retractor lentis muscle can shift the lens caudally with respect to the retina.
As in most vertebrates, the shape of the eye can be derived from laws of optics and the demand of smooth rotational movement. Although the shape can be affected by other factors (see below), for the model we shall start with a hemispherical one.
It is obvious that for proper vision the eye must have a position at the surface of the head, but its rostrocaudal and dorsoventral situation can vary in relation to constructional demands or required visual fields.
Conceiving the eye as a functional component, which is equal in shape and basic construction and has similar optical properties in the fishes compared, only differences in size and retinal composition will be considered in relation to functional differences. The latter are related to behavior and environment.
Among the ecological factors, wavelength and the amount of light stand out for observing food, partners, and enemies. As observation is distance dependent, we shall first discuss the accommodation mechanism and subsequently the structure of the retina in relation to resolution and sensitivity.
The distance from which objects are to be observed may affect the balance between demands of retinal resolution and sensitivity. Nearby objects form larger images on the retina and from them more light enters the eye than from similar objects far away. Also, accurate focusing on nearby objects asks for more precise adjustment. Therefore, catching prey items at a distance less than one cm in front of the snout demands a perfect accommodation mechanism of the lens in a rostrad-caudad direction. This is provided by the retractor lentis muscle and an antagonistic force produced by the elasticity of the lens-suspension tissue inside the eye. For such nearby vision, a high resolution is possible because in that position of the prey, the most image-forming light is available. Therefore, the highest resolution can be expected in the caudal part of the retina (sharp, myopic vision).
Retinal resolution is inversely related to the separable angle and consequently proportional to the density of photoreceptors and to eye size (Fig. 2). With the same optimal packing of receptors, and ignoring differences in neuronal convergence, sensitivity depends only on the size of the photoreceptors because the amount of photons arriving at the retina per retinal square unit and per unit time is independent of eye size (Van der Meer and Anker, 1986). Maximal packing of receptors in the retinal plane depends on their shape and configuration but not on the size of the receptors (Van der Meer, 1992). It may be noted that the receptor convergence, which is the ratio between the number of receptors and the number of ganglion cells, is positively related to sensitivity and negatively to resolution.
Per vision angle more equal receptors or the same number of larger receptors can be placed in a bigger eye (Fig. 3). This simple model formulates a possible increase of retinal resolution or an increase of sensitivity or an increase of both in bigger eyes (this indicates a trade-off relation between resolution and sensitivity). The actual solution will probably be a compromise of which the degree is related to the niche of the animal. In general, we can expect fishes feeding on large prey and detecting prey from a long distance to have large eyes with big photoreceptors, whereas fishes feeding on nearby small items will have high densities of small receptors, particularly in the caudal part of the retina. There are, however, many exceptions to these expectations. Apparently there are more ecological demands. Also, it is possible that in the retina different areas serve different functions and consequently show differences in structure. Moreover, light intensity is a very important factor. Exceptions may also be due to the limited capacity and space, so that the demands cannot be met.
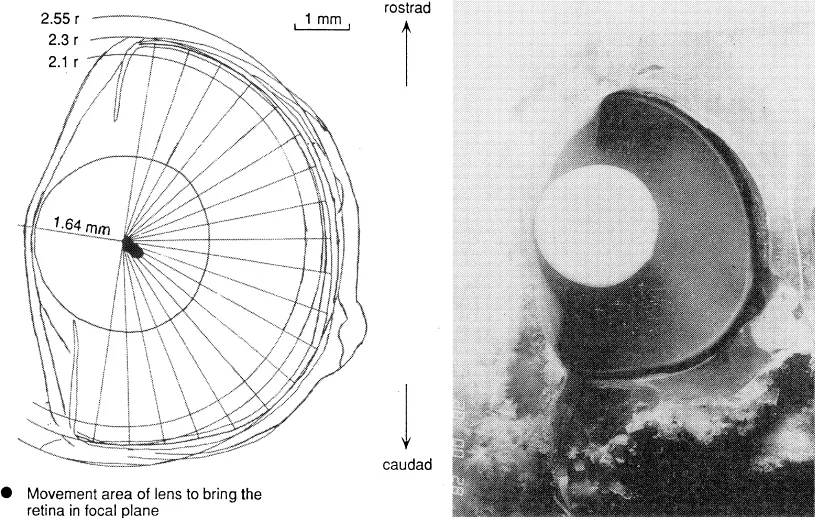
Fig. 1. Position of the lens in a horizontal section of the eye through the center of the lens.
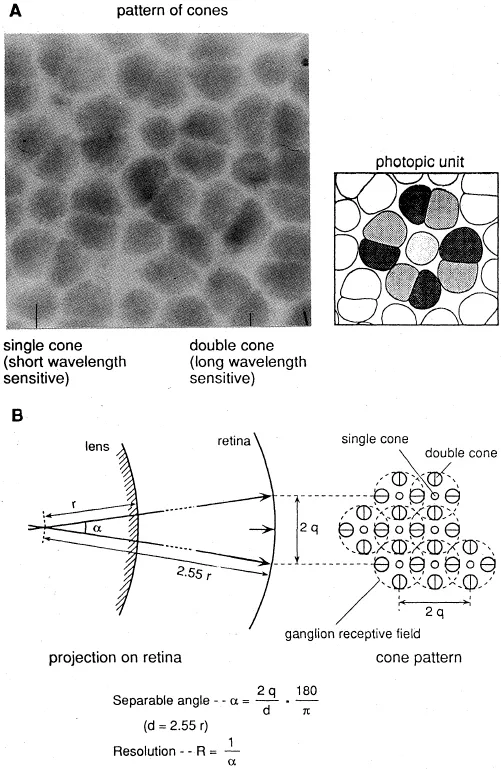
Fig. 2. A square pattern of cones and the supposed photopic unit in this pattern. В. Manner in which separable angle and resolution are calculated from the supposed ganglion receptive field (after Van der Meer and Anker, 1984).
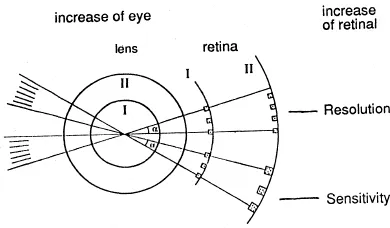
Fig. 3. Possible enhancement of retinal resolution or sensitivity (or both) when the size of the eye increases (after Van der Meer and Anker, 1986).
The following examples show some of these expectations. The fact that small fry of most species grow up in shallow coastal areas can be a consequence of the limited capacity of their small eyes for vision in darker habitats. The actual increase of resolution during ontogeny, as experimentally determined by Van der Meer (1991), appeared to be related to the size of photopic units, which were twice as small as calculated (Fig. 2). An experimentally determined increase in sensitivity was found to be related to increase in receptor volume and probably to increase in photosensitive pigment in the receptor outer segment (Van der Meer, 1991).
A predicted general increase in resolution and sensitivity related to eye size was found during ontogeny (Van der Meer, 1991) and in adults of eleven cichlid species differing in eye size (Van der Meer and Anker, 1984). Also the predicted relatively higher resolution of the temporal area of the retina was confirmed. Much variation occurred depending on the behaviorally and environmentally required balance between resolution and sensitivity.
To make the model more operational to testing, it has to be refined for specific cases for which the ecological situation is well known. It can be refined by adding the property that the single cones contain short wavelength sensitive pigment and the double cones contain long wavelength sensitivity pigments (Van der Meer and Bowmaker, 1955). If the wavelength is an important demand, it is predictable that in darker habitats those receptors should be increased in size which are most sensitive to the available wavelength and the others should be decreased to keep sufficient resolution. Ultimately the ratio in numbers between both receptor types may change.
In the turbid water of Lake Victoria, long wavelengths penetrate deeper into the water layers than shorter ones (de Beer, 1989). From these facts it can be predicated that in cichlids living in deeper water layers, single cones should decrease in size, or even be absent to accommodate larger double cones. From a model study (Van der Meer, 1992), it appears that a change from a square pattern with larger single cones to a row pattern with small or without single cones becomes necessary to maintain an optimal packing.
These predictions have been tested by breeding fish under several artificial light conditions (Van der Meer, 1993) and by comparing fishes with more or less the same feeding behavior living in optically different surroundings (Van der Meer, 1991). Indeed, in fry of Haplochromis sauvagei deprived completely of short wavelength light, a rowlike pattern with enlarged double cones and smaller single cones develops, whereas square patterns of receptors are observed in wild specimens and in specimens raised in the laboratory under normal light conditions. Such a change in pattern can also be expected when growing fry migrate to deeper water. However, retinal development of different species (Haplochromis pyrrhocephalus and H. argens) raised under similar laboratory conditions showed a different retinal configuration from the start. In general, species of deeper habitats in Lake Victoria have row patterns, whereas most species living in shallow waters have a square pattern with distinct single cones (Fig. 4).
Goldschmidt et al. (1990), made an interesting comparison of four related Hap lochromis species sampled from the same water column (14 m deep) in the Mwanza Gulf of Lake Victoria (Fig. 5). One species, H. argens, was a surface dweller to a depth of 4 m during day and night. H. heusinkveldi was found distributed over the whole column during the day and more concentrated in the upper 4 m during the night. H. pyrrhocephalus was found concentrated in the lower part of the column during the day and also moved to the upper layer during the night. H. reginus was a bottom dweller during day and night and migrated from deeper locations during its breeding period. The predicted change from a square pattern to a row is confirmed by comparing the retinas of the four species. H. argens has a square pattern with distinct single cones, H. reginus a row pattern without single cones (Fig. 4). The retinas of H. heusinkveldi and H. pyrrhocephalus have an intermediate configuration. The expected shift of balance between resolution and sensitivity in favor of sensitivity related to depth and consequently to decreased luminance of longer wavelength was not confirmed. Mean cone densities in specimens with equal eye size of these four species did not decrease with depth and no distinct decrease in mean cone size was found (Van der Meer, 1991). Only H. pyrrhocephalus has distinctly larger cones and a slighter resolution if mean values for whole retinas are compared. During the ontogeny of the four species cone density per visual angle as well as cone size increases with eye size. The intraretinal variation in the four species is much more difficult to interpret (Fig. 7).
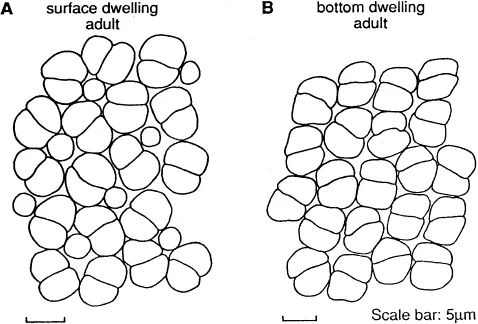
Fig. 4. A. Square pattern of double cones surrounding single cones in H. argens. B. Row pattern of double cones in H. reginus.
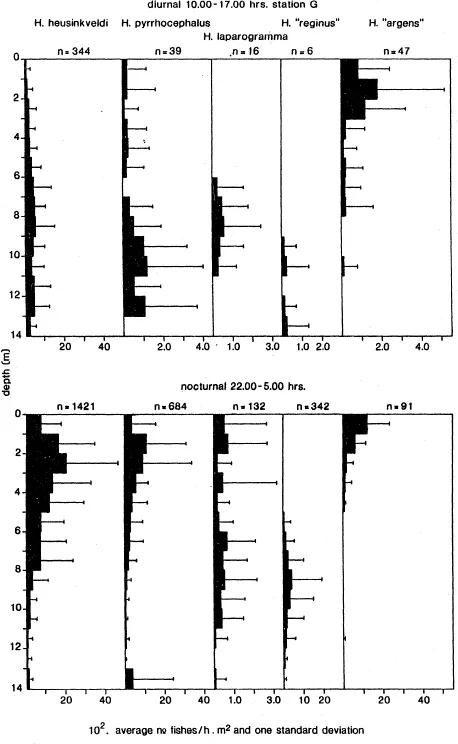
Fig. 5. Diurnal and nocturnal vertical distributions of five zooplanktivores. Average number/h.m2 of net (and one standard deviation) (abscissa) for every meter in depth (ordinate) plotted. Averages based on 33 diurnal and 43 nocturnal observations, from 1981–1984, at station G on the Mwanza Gulf transect. n = number of individuals (after Goldschmidt et al., 1990).
Hence we are compelled to look for other ecological factors, e.g. food. From the investigation of Goldschmidt et al. (1990) of the four species, and the food distribution over the water column in the period 1981–1984, it appears that (Fig. 6):
a. During the day all four species fed on Zooplankton, which was distributed over the whole column with, in the dry season, peak values in the top layers during both day and night. Besides Zooplankton, H. reginus fed also on detritus and phytoplankton, mainly diatoms (Melosira), and H. heusinkveldi also ate bluegreen algae. Except for H. reginus, insect larvae were found only occasionally in the stomach of the other species...