Neurophysiology: A Conceptual Approach offers a refreshing alternative to 'learning by rote'. Under new authorship, the sixth edition preserves the legacy of the original author, the late Roger Carpenter, retaining the concise approach and readable style so central to its predecessors. Integrating the disciplines of neurology and neuroscience with an emphasis on principles and functional concepts, this comprehensive textbook covers the entire subject of neurophysiology, from the conduction of nerve impulses to the higher functions of the brain, within a single accessible volume.
Key Features:
Everything the student of medicine or physiology needs to understand neurophysiology.
Blends successfully the principles of neuroscience with clinical manifestations in line with modern undergraduate curriculums.
Revised and updated, with a particular focus on proprioception, skin sense and hearing, including developments in cochlear implants, and functional MRI
Over 500 illustrations, accompanied by full figure legends, also available as a download for use in presentations.
Choice of PB with bundled ebook, durable HB or ebook only for complete flexibility
Full of explanatory colour diagrams, the book remains an unrivalled 'one-stop shop' for students of medicine, physiology and applied physiology, neurophysiology, neuroscience, and other bioscience disciplines seeking an integrated introduction to the challenging disciplines of neuroscience and neurology.
Häufig gestellte Fragen
Wie kann ich mein Abo kündigen?
Gehe einfach zum Kontobereich in den Einstellungen und klicke auf „Abo kündigen“ – ganz einfach. Nachdem du gekündigt hast, bleibt deine Mitgliedschaft für den verbleibenden Abozeitraum, den du bereits bezahlt hast, aktiv. Mehr Informationen hier.
(Wie) Kann ich Bücher herunterladen?
Derzeit stehen all unsere auf Mobilgeräte reagierenden ePub-Bücher zum Download über die App zur Verfügung. Die meisten unserer PDFs stehen ebenfalls zum Download bereit; wir arbeiten daran, auch die übrigen PDFs zum Download anzubieten, bei denen dies aktuell noch nicht möglich ist. Weitere Informationen hier.
Welcher Unterschied besteht bei den Preisen zwischen den Aboplänen?
Mit beiden Aboplänen erhältst du vollen Zugang zur Bibliothek und allen Funktionen von Perlego. Die einzigen Unterschiede bestehen im Preis und dem Abozeitraum: Mit dem Jahresabo sparst du auf 12 Monate gerechnet im Vergleich zum Monatsabo rund 30 %.
Was ist Perlego?
Wir sind ein Online-Abodienst für Lehrbücher, bei dem du für weniger als den Preis eines einzelnen Buches pro Monat Zugang zu einer ganzen Online-Bibliothek erhältst. Mit über 1 Million Büchern zu über 1.000 verschiedenen Themen haben wir bestimmt alles, was du brauchst! Weitere Informationen hier.
Unterstützt Perlego Text-zu-Sprache?
Achte auf das Symbol zum Vorlesen in deinem nächsten Buch, um zu sehen, ob du es dir auch anhören kannst. Bei diesem Tool wird dir Text laut vorgelesen, wobei der Text beim Vorlesen auch grafisch hervorgehoben wird. Du kannst das Vorlesen jederzeit anhalten, beschleunigen und verlangsamen. Weitere Informationen hier.
Ist Carpenter's Neurophysiology als Online-PDF/ePub verfügbar?
Ja, du hast Zugang zu Carpenter's Neurophysiology von Dunecan Massey, Nick Cunniffe, Imran Noorani im PDF- und/oder ePub-Format sowie zu anderen beliebten Büchern aus Medizin & Anatomie. Aus unserem Katalog stehen dir über 1 Million Bücher zur Verfügung.
This book is about trying to understand the brain; we might begin by asking whether such an aim is not, in fact, hopelessly ambitious. Your brain is a machine whose complexity far exceeds anything constructed by human hands. It is made up of units — cells called neurons — that provide both the pathways by which information is transmitted within the brain and also the computing machinery that makes it work. There are quite a lot of these neurons: about thirty times as many as the entire population of this planet. Another not very useful fact to impress your friends is that every cubic inch of your cerebral cortex has 10 000 miles of nerve fibre in it. A typical neuron is wired up to a thousand or two of its neighbours, and it is the pattern of these connections that determines what the brain does. All this makes the brain a fantastically complex structure, a thicket of twisting, interweaving fibres.
Our brains need to be complex: part of what they do is to embody a kind of working model of the outside world, that enables us to imagine in advance what will be the result of different courses of action. It follows that the brain must be at least as complicated as the world we experience. So the study of the brain is like the study of human society: for a society can only be fully understood if we comprehend not just the behaviour of isolated individuals, but also the interactions that each of them makes with others. Understanding the brain is a task as daunting as trying to comprehend the behaviour of the entire human race, its politics, its economics, and all other aspects of what it does; in fact, about thirty times more difficult. As a result, the study of the brain has in some respects a closer affinity with ‘arts’ subjects like history than it does with much conventional science. For many people that is its attraction.1
The need for neurons
How has such complexity come about? The evolutionary history of the brain is not well understood but in broad outline was certainly something like the account that follows.
For a single-celled organism such as an amoeba, coordination is essentially chemical: its brain is its nucleus, acting in conjunction with its other organelles.2
But a multicellular organism clearly needs some system of communication between its cells, particularly when, as in the primitive invertebrate Hydra, they are specialised into different functions: secretion, movement, nutrition, defence and so on. Communication between cells practically always means chemical communication. One cell releases a chemical substance; somewhere else another cell is waiting for this chemical, which tells it to do something. Nearly always, the target cell has receptor proteins in its cell membrane that recognise the particular chemical. Often these form part of what are technically called ligand-gated channels in the membrane, opening in response to the chemical and often allowing ions such as calcium, which act as messengers, to enter the cell. This kind of chemical communication is most familiar as hormones, operating within the body, perhaps to cause widespread changes: a good example is adrenaline, released in dramatic circumstances and alerting more or less the whole body. The chemical is not directed at any particular cell: it is broadcast throughout the whole body.
Diffusion time
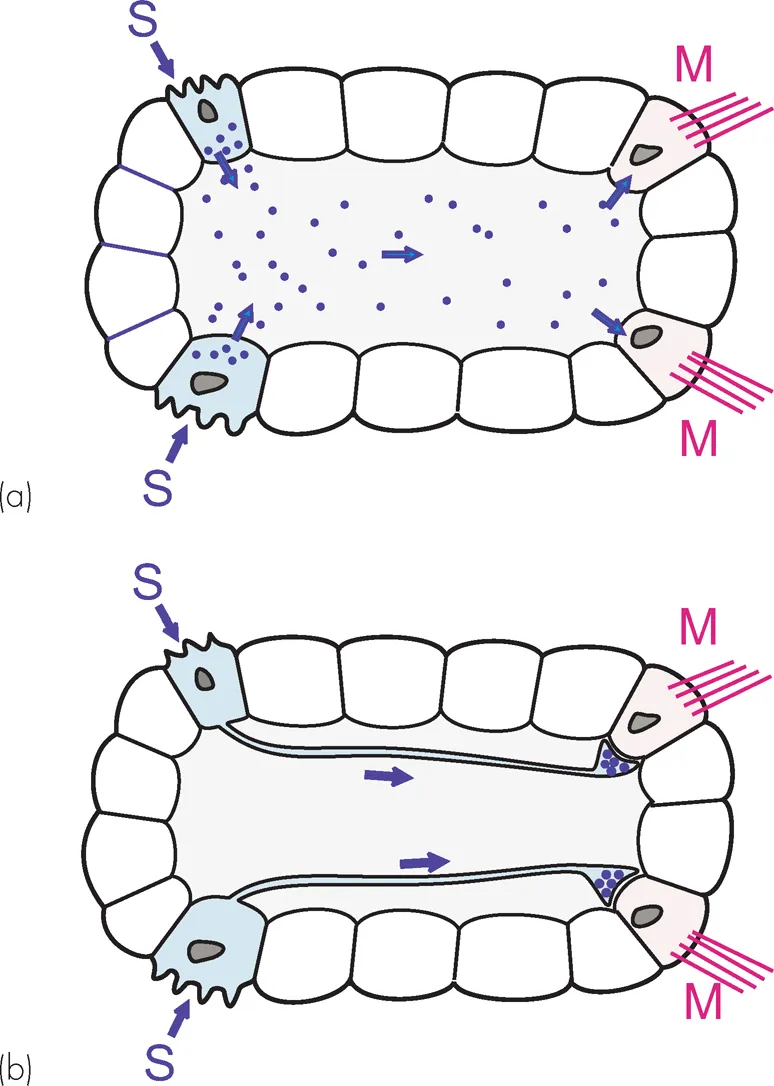
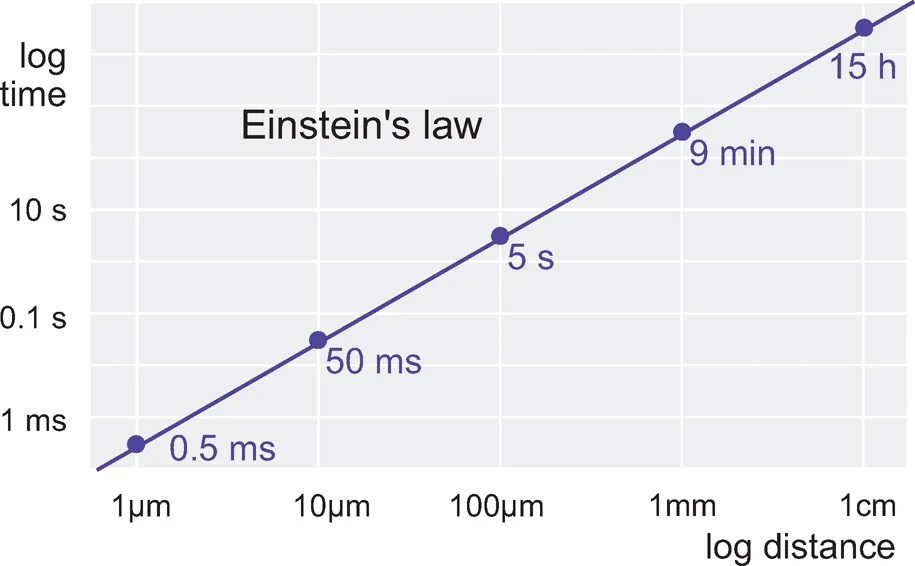
In small and slow creatures, this works fine. The one shown in Figure 1.1 has sensory cells (S) that control motor cells (M) by releasing a chemical transmitter or hormone into the common fluid space. But as an organism gets bigger, things get difficult, in two ways. First of all time: how long it takes for the message to get from the sender cell to its recipient depends on diffusion. It depends rather dramatically, because diffusion time is proportional to the square of the distance travelled — Einstein’s Law (Fig. 1.2) — so it takes a disproportionately longer time for a chemical signal to travel over increasing distances: 1 tm takes 0.5 ms, but 10 mm takes 15 hours. If speed of response is not particularly important, this kind of communication may still be satisfactory even in very large organisms: our own hormonal control systems are of course precisely of this kind. Circulation helps, but even so your blood takes about a minute to go once round the body. If a tiger suddenly burst into your room, although you would instantly release a burst of adrenaline into your bloodstream, it would take about a minute for it to reach its target tissues. By that time you might well have been devoured.
Figure1.1 (a) A hypothetical multicellular organism with sensory cells (S) that control motor cells (M) by releasing a chemical transmitter or hormone into the common fluid space. (b) Direct connections between sensory and motor cells by means of nerve axons, providing communication that is both far quicker and more specific. Their ultimate action on the motor cells is still chemical.
Specificity
The other problem is specificity: the little creature in Figure 1.1 can do just two things, but we can do much more. Each of us has well over 1200 separate muscles, for a start. And as we get more complicated with lots of different receptor cells and lots of different muscle cells, we want to do more specific things in response to specific circumstances. In a hormonal system, specificity can only come about by having a range of different chemicals, with target cells responsive to some but not others. In fact there is a very long list of hormones in the body, running into hundreds — but still not enough for the countless actions we might want to undertake. What is the solution?
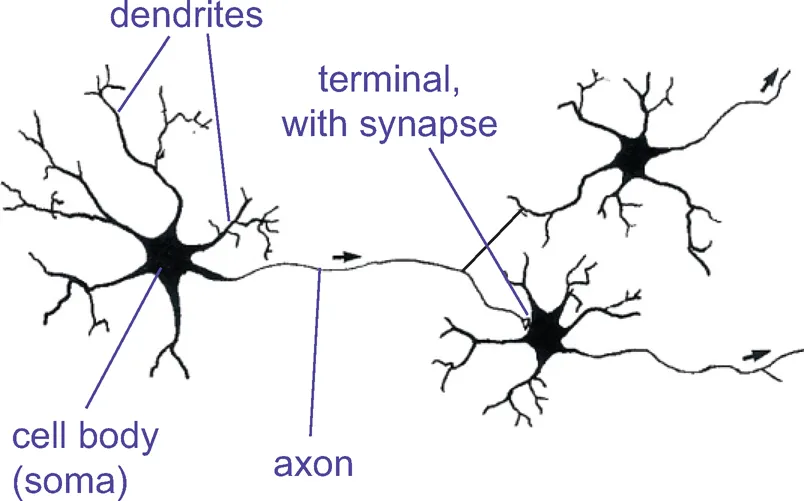
In hormonal systems, a message is, in effect, shouted throughout the whole body. One way of being more specific is, as it were, to whisper it confidentially into the ear of the target cell. And this precision is where neurons come in. Neurons are simply cells with special elongated output processes called axons, which can often be very long indeed — the longest neurons measure about a metre long. The axons grope out towards their targets and make physical contact with them at regions called synapses (Fig. 1.3). They still release their chemical at the synapse, but this time it only affects the cells it makes contact with and not the others, so we no longer need lots and lots of different transmitters. For instance, every muscle in your body is controlled by the same chemical substance, acetylcholine. Though specific, neurons can still be widespread in their actions: because axons can branch, they are capable of influencing thousands of target cells that can be scattered over a very wide area. These axons end either on muscles (where the synapse is called a neuromuscular junction), on the cell body (soma) of a target neuron, or often on the tree-like dendrites that enormously increase the surface area of many neurons.
Figure1.2 Diffusion time is a function of distance: note the logarithmic scales.
It is still chemical communication, but we call the chemicals transmitters rather than hormones. There are several dozen known transmitter substances, falling into three main groups: amino acids, amines and peptides. In fact, knowing the name of a transmitter doesn’t tell you much, because transmitters don’t have to do the same thing at every site. The receptor molecules on the target cell translate arrival of the transmitter into some specific action, but different cells may use different receptor proteins and therefore translate the event differently. Thus there is no necessary logical connection between what the transmitter is and what it does. For example, acetylcholine excites skeletal muscles but inhibits heart muscle: the receptor proteins in the heart muscle cell membrane are different from those in skeletal muscle. The message is the same, but the meaning is different. So nerves provide a system that is very specific both in what it does and where it does it. But what about speed? As we have seen, diffusion down long neurons would be hopelessly slow for distances of more than a few microns. Yet the fastest neurons can actually convey information at over 100 m/s. They are able to do this because they use a much quicker physical process than diffusion, namely electricity and the flow of current: these processes are discussed in the next chapter.
Figure1.3 Schematic representation of a ‘classical central neuron synapsing with two others, in one case on a dendrite and in the other on the cell body Or soma.
Clinical box 1.1 Neurotransmitters as drug targets
The issue of specificity is one of the first hurdles facing the clinician. As noted above, there are only a limited number of transmitter chemicals to bear a near infinite number of bits of information. The evolutionary solution has been the neuron, giving precision to the signal using spatial coding by delivering the transmitter to a particular recipient. Rather as the order ‘fire’ is one of a limited number of commands a sniper might expect to hear in the field, the general accomplishes specificity by ensuring the command is issued to the correct person; confusing his or her cables could clearly deliver a suboptimal result. The problem the clinician faces when trying to restore brain function by using administered analogues of neurotransmitters is that of targeting the signal to the correct recipient.
For example, morphine acts on a family of receptors and mimics endogenously generated signals called opioids. We know morphine as a rather powerful painkiller, and, not surprisingly, it acts on neurons that block pain signalling. Unfortunately, there are other morphine receptors located elsewhere in the body, the activation of which has rather less desirable characteristics; for example, neurons exist in ganglia near the bowel, which, when activated by morphine, reduce gut motility. Indeed, one of the most distressing side effects of morphine is constipation. The body gets around this problem by targeting its opioid release to those si...