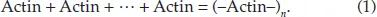![]()
PART I
Theoretical Approaches
![]()
1. Molecular Biocommunication
Alexei A. Sharov
Laboratory of Genetics and Genomics
National Institute on Aging251 Bayview Blvd., Baltimore, MD 21224, USA
[email protected]
The integrity of living cells is maintained via regular communication between cell components. Membrane receptors provide information on the environment and launch signals to modify cellular functions accordingly. Gene regulatory networks coordinate the work of genes to keep the homeostasis of the cell and optimize its functions. Ribosomes are miniature robots that are programed to produce various kinds of proteins. Molecular biocommunication includes the most primitive form of sign processes in nature (I call it protosemiosis), where signs directly encode and control cell functions instead of being associated with objects. Such signs can be classified into proto-icons that operate via specific binding, protoindexes that signal via physical association of several binding and/or catalytic domains, and proto-symbols that are processed by universal subagents equipped with a set of heritable adapters. The functions of molecular signaling include fabrication of cell components, sensing, memory, movement, and propagation of depolarization waves on membranes. Besides immediate cellular functions, molecular networks support robustness, error control, and modularity, which often require categorization of objects, and therefore belong to the advanced type of sign processing — eusemiosis.
1. Introduction
The science of communication gradually expanded with the advance of human technology and knowledge. It was applied first to human language, then, subsequently, to technology, animal acoustic, and chemical signals. With the emergence of molecular biology, the field of communication had unexpectedly expanded to the level of molecules within cells. Molecules play various roles in biocommunication. Membrane receptors provide information on the environment and launch signals to modify cellular functions. Cytoplasmic and nuclear receptors coordinate the work of genes to keep the homeostasis within cells and optimize their functions. Ribosomes are miniature robots that are programed to produce various kinds of proteins. Heredity is based on the long-term memory molecules [e.g., deoxyribo nucleic acid (DNA) and ribonucleic acid (RNA)] which are copied and passed from one generation to another. Molecular communication ensures the functional integrity of cells, and supports their growth, reproduction, and defense from pathogens. The study of communication became an interdisciplinary field where the original linguistic terminology is not fully applicable. One of the first attempts to overcome the constraints of linguistics was undertaken by Charles Peirce who developed a general theory of signs (Peirce, 1998). He defined signs as follows: “a sign is something which stands to somebody for something in some respect or capacity” (Peirce, 1931: 228). Activities of living organisms associated with sign perception, interpretation, or creation are called “semiosis”. Peirce considered mind as an important condition for the existence of signs: “The third condition of the existence of a sign is that it shall address itself to the mind. It is not enough that it should be in relation to its object […] but must be regarded by mind as having that relation” (Peirce, 1986: 83). But he thought that sign relationships can be captured by any mind, and thus they are observer independent. Thus, causal relationships or physical connections are sufficient to make a sign: “if I take all the things which have certain qualities and physically connect them with another series of things, each to each, they become fit to be signs” (Peirce, 1998: 40). However, he did not envision (as anyone else at that time) that communication may exist between mindless molecular agents that are not capable of contemplating objects. Thus, the analysis of molecular biocommunication requires more general approaches than Peirce’s semiotics.
In this chapter, I argue that molecular biocommunication includes the most primitive form of sign processes in nature, which I call “protosemiosis”, a term that was earlier suggested by Prodi (1988), where signs (proto-signs) are not associated with objects, but instead directly control the activity of agents (Sharov, 2013; Sharov and Vehkavaara, 2015). The terms “control” and “regulation” are interpreted by some authors as efficient causation or brute force (Hoffmeyer and Emmeche, 1991; Bruni, 2008). In contrast, by “control” I mean various kinds of effects which do not fall into the category of efficient causation: initiation or inhibition of actions, specifying the mode of action, its direction, timing, and scope, as well as conditional (i.e., context-dependent) effects. Although proto-signs contribute to the actions, they never fully determine them because actions are always agent dependent and agent-specific. Molecular proto-signs are classified here into proto-icons, proto-indexes, and proto-symbols. Higher levels of molecular communications support error correction, robustness, and modularity of signaling, as well as lasting interactions with internal and external objects. Corresponding molecular networks often include internal representations of objects as fixed point attractors in the continuous signaling phase space (i.e., categorization). These belong to the advanced type of semiosis, I call it “eusemiosis”, where signs are associated with objects or object states. Analysis of molecular communication is a part of a wider endeavor, biosemiotics, which is aimed at the integration of semiotics with biology at all levels of organization (Hoffmeyer, 2008).
2. Chemistry Versus Semiotics: How Does a Molecule Become a Sign?
Biophysics and chemistry considers a living cell as a complex chemical reactor, where each process can be described in terms of reactants, products, catalysis, and kinetics. However, chemistry tells nothing about the purpose of chemical interactions, and thus, it is not sufficient to understand how the cell works. Molecular interactions in living organisms should be studied not only by chemistry but also by biosemiotics which attempts to answer how biological processes are encoded, controlled, and communicated, and how do they contribute to the goals of living systems (Sharov, 1992, 2010).
Application of semiotic terms to molecular interactions raises an important question: where is a boundary between chemistry and semiotics and how does a molecule become a sign (Pattee, 1969)? Sign theory is often built on the basis of “semiotic realism”, where signs are viewed as real and objective natural phenomena without a reference to interpreters (Vehkavaara, 2003). For example, formal and final causations are used as criteria of sign relationships (Deely, 1994; Bruni, 2008). Another common criterion of a sign relationship is a source of energy: semiotic interactions are powered by the internal energy of the receiving system rather than by the impact (Bateson, 1972). However, these approaches are meaningful only if applied to goal-directed agents that use forms to produce tools and resources. Attempts to expand formal and final causations beyond agents ignore the striking difference between living and non-living systems. In particular, information was associated with purely physical categories such as entropy, negentropy (Brillouin, 1953), or with quantum physics (Green, 2000). Another theory extended quantum principles to information energy complementarity (Ji, 2012). As a result, either life is reduced to non-equilibrium physical systems (Chaisson, 2005) or life properties are extended to the full universe as in pansemiosis, hylotheism, or the Shillongator model (Merrell, 1996; Brier, 2008; Ji, 1991). The energy criterion of signs also does not work without agency: for example, a nuclear fission reaction is based on the release of internal nuclear energy, but it is not a semiotic process.
A short answer to the question posited in this section is that physics and chemistry study things as they are, whereas semiotics studies things as they are used by agents (e.g., living organisms and man-made automata) to encode and control their activities (Sharov, 2010). In other words, semiotics is focused on the role of things in the context of agent activities. Agents can be studied by physics (e.g., biophysics), but only in non-semiotic aspects (i.e., excluding goal-directed behavior and evolution). In particular, information is not a chemical or physical concept (Emmeche, 1999). In contrast, semiotics offers a comprehensive approach to the analysis of agents and signs. This criterion for separating biosemiotics from physics resembles Cartesian dualism, however, there are important differences. First, Cartesian dualism opposes material and mental substances, and the mental world is assumed to be non-material and non-physical. In biosemiotics, agents are material and their change does not contradict physics but is not fully predicted by physics. In simple living organisms, goal-directed activities are explained by the “natural self-interest”, which emerges from recursive self-maintenance and self-reproduction of agents (Vehkavaara, 2003). In addition, advanced organisms develop a network of secondary goals and habits which are partially independent from the basic natural self-interest. The attribute “being an agent” is not physical because it is impossible to make a universal detector of agency (Sharov, 2010). To identify a thing as an agent we have to understand its goals, functions, origins, external and internal communication, and these characteristics cannot be measured without a prior knowledge for the functional organization of each kind of agents. Thus, identification of agents is always non-trivial and requires a competent interpreter, which is an agent itself. Agents were often misidentified in the history of human civilization. For example, celestial bodies were considered as super agents (gods), but infections were not recognized as micro-agents. Moreover, nobody suspected that our body is made of trillions of tiny subagents called cells. Second, agents differ in their functional complexity, and agents at the lower end of the complexity scale do not possess mind. Thus the distinction between agents and nonagents is different from the matter/mind dichotomy in Cartesian dualism. Third, biosemiotics considers a transition between nonagents and simple agents at the origin of life; in contrast, mind did not originate from matter according to the Cartesian doctrine.
The difference between chemical and semiotic approaches can be illustrated by the reaction of actin polymerization. Actin is a very abundant protein in eukaryotic cells which makes a cytoskeleton via polymerization into long fibers. Actin molecules emerged in evolution for a specific purpose: to make flexible filaments that can be easily assembled, disassembled, and integrated into the cytoskeleton of cells. These filaments increased cell stability, preserved its shape, and allowed crawling (Lodish et al., 2000). Large eukaryotic cells could never have appeared in evolution without a cytoskeleton. Polymerization of actin can be viewed as a simple chemical reaction:
Actin protein is polarized and its (+) end fits due to its shape and charge with the (–) end of another molecule. Thus, actin monomers can polymerize into fibers in solutions with sufficient ionic strength (Lodish et al., 2000). This reaction, however, also represents a simple case of signaling. Actin monomers readily bind to the terminal actin molecules in the fiber but not to free-flowing actins. This means that attachment of actin molecule to the filament makes a signal for binding of the next molecule. Thus, the signal travels along the filament at the tip of its growth and results in the construction of the full filament.
Polymerization of actin is controlled by additional signaling molecules. The actin monomer has a site for binding adenosine triphosphate (ATP) (along with Mg++), which facilitates polymerization at the (+) end of the filament (Lodish et al., 2000). However, the ATP is hydrolyzed to adenosine diphosphate (ADP) immediately after incorporation of monomers into the filament (Fig. 1). Dissociation of actin occurs at the (–) end, which results in the “treadmill effect”. Tropomodulin, however, caps actin polymers at the (–) end and prevents the dissociation. Another protein, CapZ, which was found in Z bands of sar...