![]()
Section 1
Structure and function
![]()
1
The cardiovascular system
Marina Sawdon
Introduction
The purpose of the cardiovascular system is to deliver oxygen (O2) and nutrients to the metabolising tissues, and carry waste products such as carbon dioxide (CO2) and heat away. The heart is made up of individual muscle cells called cardiac myocytes, arranged to form a functional syncytium (acting as one muscle) which allows an electrical signal to spread across the heart in a coordinated way. This coordinated electrical activity leads to the mechanical beat; pressure is generated in the walls of the heart to eject blood from its chambers, to be delivered via the blood vessels to the lungs and systemic tissues.
Anatomy of the cardiovascular system
Gross anatomy of the heart
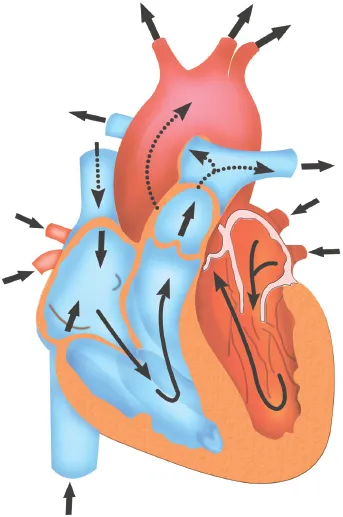
The cardiovascular system is functionally two circulations arranged in series: systemic and pulmonary (blood is pumped from one circulation into the other). The heart is structurally and functionally two pumps. The right side pumps blood into the pulmonary circulation to allow oxygenation of the blood in the pulmonary capillaries (see Chapter 3 on the respiratory system). The left side of the heart pumps blood into the systemic circulation to deliver that oxygenated blood to the tissues, and carry waste products (such as CO2) back to the right side of the heart and thus the pulmonary circulation for excretion (see Figure 1.1). In a normal, healthy, adult heart, there is no direct connection between the two.
Chambers
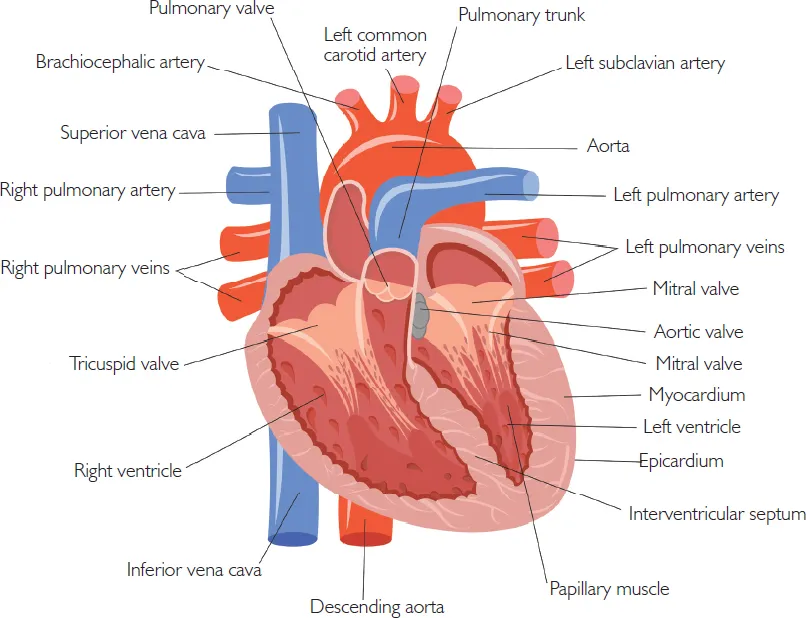
The heart is made up of four chambers: two atria and two ventricles (one of each on each side of the heart). The right ventricle pumps blood via the pulmonary artery into the pulmonary circulation. Oxygenated blood returns from the pulmonary circulation back to the left atrium via the pulmonary vein. Blood from the left atrium drains into the left ventricle down a pressure gradient, and – from there – is ejected into the systemic circulation via the aorta. Oxygenated blood is delivered to systemic tissues via arteries, arterioles and capillaries (see the section on the structure and function of the blood vessels, p. 5). Deoxygenated blood then returns back to the heart via venules and veins, and passively drains into the right atrium via the vena cavae.
Figure 1.1 The direction of blood flow through the heart
Valves
Blood is allowed to flow in one direction through the heart due to strategically placed one-way valves (see Figure 1.2). Blood flows from the atria into the ventricles via the atrioventricular valves (the tricuspid valve on the right side of the heart and the mitral valve on the left). Blood leaves the ventricles and enters the arterial system via another set of valves; the semilunar valves (the pulmonary valve on the right side of the heart and the aortic valve on the left).
Figure 1.2 The heart valves
Structure and function of the blood vessels
The heart generates pressure to help drive the blood around the circulation to deliver oxygen to the tissues, and deliver deoxygenated blood back to the heart. This pressure acts as a unit of energy and the pressure is therefore used up (rather like petrol being used to drive a car from A to B) to move the blood around the circulation. By the time the blood enters the veins to return back to the heart, the blood pressure is extremely low.
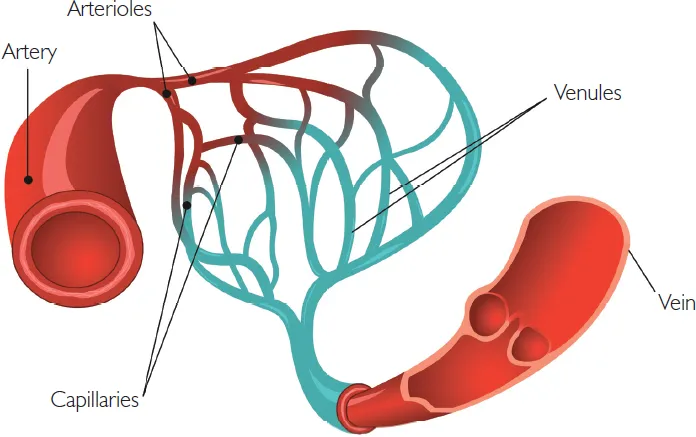
Blood vessels have some common features, but also some differences that are related to their particular function. All blood vessels are lined with a single layer of endothelial cells, called the tunica intima. In arteries and veins, the layer of smooth muscle cells covering the tunica intima is called the tunica media; and the thin outer layer of connective tissue in arteries, arterioles and veins is called the tunica adventitia and gives these vessels some stability.
The structure of each class of blood vessel is related to its function.
Arteries
Arteries deliver lots of blood, very fast and under high pressure, to the arterioles. They are therefore large in diameter, and have lots of elastin and smooth muscle around their walls (see Figure 1.3). These thick muscular walls help the arteries resist collapse.
Arterioles
Arterioles also have lots of smooth muscle but this smooth muscle is highly innervated by the sympathetic nervous system. Activity in the sympathetic nervous system leads to changes in luminal diameter and hence resistance to blood flow (see the section on regulation of blood pressure; the baroreceptor reflex, p. 12). This regulates the flow rate downstream into the tissue capillaries (helping to match blood flow, and hence oxygen delivery to the needs of the tissue), and also helps regulate systemic blood pressure upstream (see the section on regulation of blood pressure; the baroreceptor reflex, p. 12). These vessels are referred to as resistance vessels because they are the main site of vascular resistance to blood flow.
Capillaries
Capillaries are the site where fluid and blood gases (oxygen and carbon dioxide) are exchanged between blood and tissues, so these vessels need to be thin-walled. They are therefore composed of one layer of endothelial cells (tunica intima) and its basal lamina, with no elastin or smooth muscle. Once blood has passed through the tissue capillaries, it becomes venous in nature.
Venules and veins
Venules and veins deliver blood back to the right side of the heart. Venules and veins are thin-walled vessels which have some smooth muscle associated with them. By the time the blood has reached the veins, pressure has fallen very low. One-way valves are therefore needed to help the blood move in one direction back to the heart.
The smooth muscles around the walls of the veins are innervated by the sympathetic nervous system. As the diameter of the veins can be quite large, they also act as capacitance vessels, and blood within them is referred to as the venous reservoir. (The veins contain approximately 60% of the total blood volume at any one time.) This blood can be mobilised in times of emergency (such as haemorrhage) through increased activity in the sympathetic nervous system, which decreases the diameter of the lumen and boosts venous return to the heart.
Microcirculation
The microcirculation consists of the arterioles (the site of resistance to blood flow), the capillaries (the site of exchange between tissue and blood) and post-capillary venules. The microcirculation is responsible for regulating blood flow to tissues, blood pressure and tissue fluid.
Figure 1.3 Structure and function of the systemic blood vessels
The relationship between pressure, flow and resistance
The relationship between pressure, flow and resistance can be described by a modified version of Ohm’s law. Ohm’s law states that an electrical difference (voltage) between two points is generated when ions flow (current) against a resistance:
V (voltage) = I (current) x R (resistance).
The heart makes blood flow against a resistance which generates a pressure. The haemodynamic version of Ohm’s law is thus:
P (pressure) = F (flow) x R (resistance).
This pressure drives blood through the vascular beds. Thus, we can rearrange the equation to regulate blood flow to individual organs:
F = P/R.
Therefore, if either blood pressure or vascular resistance changes, blood flow to the tissue beds will change. Globally, the haemodynamic version of Ohm’s law is:
Systemic blood pressure = cardiac output x total peripheral resistance
(BP = CO x TPR).
The regulation of systemic blood pressure will be covered later in the section on regulation of blood pressure; the baroreceptor reflex (p. 12).
Cardiac output = heart rate x stroke volume (CO = HR x SV).
The regulation of cardiac output will be covered later in the section on regulation of cardiac output (p. 10).
The origin of the heartbeat
The heart is made up of individual cardiac muscle cells called myocytes. For the heart to function efficiently as a pump, these individual cells must beat in a coordinated fashion. The coordination of the heartbeat originates with the coordinated spread of electrical activity across the myocytes. This electrical activity leads to the mechanical beat.
Electrical activity
It is important to have an understanding of the electrical activity of the cardiac myocytes, not only to understand the electrocardiogram (ECG), but also to understand how and where drugs act that are used to treat conditions such as rhythm disturbances.
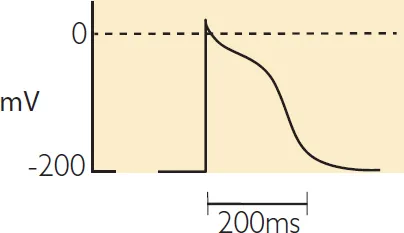
A membrane potential is the electrical difference (voltage) across the cell membrane due to the difference in the distribution of anions (negatively charged ions) and cations (positively charged ions). By convention, the extracellular fluid is considered to be at zero volts and so all electrical potentials are relative to this. There is a potential (electrical difference) between the inside and the outside of the cell such that inside is negative with respect to the outside, at rest (i.e. when no action potential is being propagated). This is the resting membrane potential. If ions are allowed to flow across the membrane, by opening and closing specific ion channels, this will alter the electrical potential of the membrane (the membrane potential) over time and trigger an action potential (see Figure 1.4). The action potential in a cardiac myocyte triggers a series of intracellular events, ultimately resulting in contraction of the cardiac muscle fibre.
Figure 1.4 An action potential
The resting membrane potential
Ions (charged particles, such as sodium, calcium and potassium) can only cross the cell membrane when their specific channel is open. Due to the concentration differenc...