
eBook - ePub
C4 Plant Biology
Rowan F. Sage, Russell K. Monson, Rowan F. Sage, Russell K. Monson
This is a test
Partager le livre
- 596 pages
- English
- ePUB (adapté aux mobiles)
- Disponible sur iOS et Android
eBook - ePub
C4 Plant Biology
Rowan F. Sage, Russell K. Monson, Rowan F. Sage, Russell K. Monson
Détails du livre
Aperçu du livre
Table des matières
Citations
À propos de ce livre
Due to many issues related to long-term carbon dynamics, an improved understanding of the biology of C4 photosynthesis is required by more than the traditional audience of crop scientists, plant physiologists, and plant ecologists. This work synthesizes the latest developments in C4 biochemistry, physiology, systematics, and ecology. The book concludes with chapters discussing the role of C4 plants in the future development of the biosphere, particularly their interactive effects on soil, hydrological, and atmospheric processes.
Foire aux questions
Comment puis-je résilier mon abonnement ?
Il vous suffit de vous rendre dans la section compte dans paramètres et de cliquer sur « Résilier l’abonnement ». C’est aussi simple que cela ! Une fois que vous aurez résilié votre abonnement, il restera actif pour le reste de la période pour laquelle vous avez payé. Découvrez-en plus ici.
Puis-je / comment puis-je télécharger des livres ?
Pour le moment, tous nos livres en format ePub adaptés aux mobiles peuvent être téléchargés via l’application. La plupart de nos PDF sont également disponibles en téléchargement et les autres seront téléchargeables très prochainement. Découvrez-en plus ici.
Quelle est la différence entre les formules tarifaires ?
Les deux abonnements vous donnent un accès complet à la bibliothèque et à toutes les fonctionnalités de Perlego. Les seules différences sont les tarifs ainsi que la période d’abonnement : avec l’abonnement annuel, vous économiserez environ 30 % par rapport à 12 mois d’abonnement mensuel.
Qu’est-ce que Perlego ?
Nous sommes un service d’abonnement à des ouvrages universitaires en ligne, où vous pouvez accéder à toute une bibliothèque pour un prix inférieur à celui d’un seul livre par mois. Avec plus d’un million de livres sur plus de 1 000 sujets, nous avons ce qu’il vous faut ! Découvrez-en plus ici.
Prenez-vous en charge la synthèse vocale ?
Recherchez le symbole Écouter sur votre prochain livre pour voir si vous pouvez l’écouter. L’outil Écouter lit le texte à haute voix pour vous, en surlignant le passage qui est en cours de lecture. Vous pouvez le mettre sur pause, l’accélérer ou le ralentir. Découvrez-en plus ici.
Est-ce que C4 Plant Biology est un PDF/ePUB en ligne ?
Oui, vous pouvez accéder à C4 Plant Biology par Rowan F. Sage, Russell K. Monson, Rowan F. Sage, Russell K. Monson en format PDF et/ou ePUB ainsi qu’à d’autres livres populaires dans Biowissenschaften et Ökologie. Nous disposons de plus d’un million d’ouvrages à découvrir dans notre catalogue.
Informations
Sujet
BiowissenschaftenSous-sujet
ÖkologiePart I
Perspectives
1
Why C4 Photosynthesis?
Rowan F. Sage
I Introduction
The net acquisition of carbon by photosynthetic organisms is catalyzed by ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) through the carboxylation of RuBP, forming two phosphoglyceric acid (PGA) molecules. In addition to RuBP carboxylation, Rubisco catalyzes a second reaction, the oxygenation of RuBP, producing phosphoglycolate (Fig. 1). Oxygenation is considered a wasteful side reaction of Rubisco because it uses active sites that otherwise would be used for carboxylation, it consumes RuBP, and the recovery of carbon in phosphogylcolate consumes ATP and reducing equivalents while releasing previously fixed CO2 (Sharkey, 1985). Oxygenation may be inevitable, given similarities in the reaction sequence for oxygenation and carboxylation of RuBP (Andrews et al., 1987). From an evolutionary standpoint, oxygenation could be considered a design flaw that reduces Rubisco performance under a specific set of conditions. Should those conditions ever arise and persist, then opportunities may exist for alternative physiological modes to evolve. C4 photosynthesis appears to be one such alternative, appearing in response to the prehistoric advent of atmospheric conditions that allowed for significant oxygenase activity and photorespiration (Ehleringer et al., 1991).
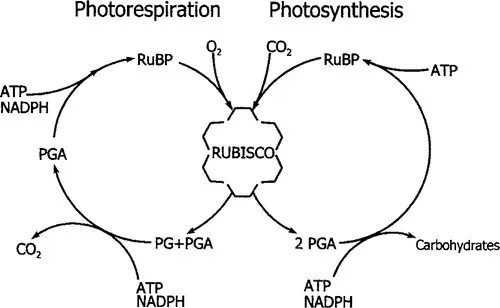
Figure 1 A schematic of the photorespiratory cycle and photosynthesis. Photosynthesis occurs when RuBP is carboxylated by Rubisco, and the products (two phosphoglyceric acid molecules; PGA) are processed into carbohydrates and used to regenerate RuBP in reaction sequences requiring ATP and NADPH. Photorespiration begins with the oxygenation of RuBP to form one phosphoglycolate (PG) and PGA, in a side reaction catalyzed by Rubisco. Processing the phosphoglycolate to PGA and eventually RuBP requires ATP and reducing power (indicated by NADPH).
II The Problem with Rubisco
Rubisco evolved early in the history of life, more than 3 billion years ago (Hayes, 1994). The CO2 content of the atmosphere at this time was orders of magnitude greater than now, and O2 was rare (Fig. 2A; Kasting, 1987; Holland, 1994). In this environment, oxygenase activity was uncommon, probably less than one oxygenation per billion carboxylations (Fig. 2C). Atmospheric O2 level remained low and was unable to support an oxygenation rate that was more than 1% of the carboxylation rate until approximately 2 billion years ago. At this time, atmospheric O2 began to rise, eventually surpassing 200 mbar (20%) about 0.6 billion years ago (Kasting, 1987; Berner and Canfield, 1989; Holland, 1994). Atmospheric CO2 level continuously declined prior to 1 billion years ago, yet at the advent of the first land plants some 450 million years ago, atmospheric CO2 was still high enough to saturate Rubisco and minimize oxygenase activity (Fig. 2C). Coal-forming forests of the Carboniferous period (360 to 280 million years ago) contributed to a reduction in CO2 partial pressures to less than 500 mbar and a rise in O2 partial pressures to more than 300 mbar (Berner and Canfield, 1989; Berner, 1994). As a consequence, Rubisco oxygenase activity is predicted to have become significant (>20% of the carboxylation rate at 30°C) for the first time in nonstressed plants at the prevailing atmospheric conditions (Fig. 2B,D). After the Carboniferous period, CO2 levels are modeled to have risen to more than five times current levels for about 200 million years, and the rate of RuBP oxygenation was again a small percentage of the carboxylation rate. Over the past 100 million years, atmospheric CO2 levels declined from more than 1,000 μbar, eventually falling below 200 μbar during the Pleistocene epoch (2 to 0.01 million years ago). In the last 15 million years, Rubisco oxygenase activity is modeled to have risen above 20% of carboxylase activity at 30°C, eventually surpassing 40% of carboxylase activity at the low CO2 levels (180 μbar) experienced during the late Pleistocene (Fig. 2D). It is only after atmospheric CO2 levels are low enough to allow the rate of RuBP oxygenation to exceed 20% to 30% of the carboxylation potential that C4 plants appear in the fossil record (Cerling, Chapter 13). No evidence exists for C4 photosynthesis during the Carboniferous (Cerling, Chapter 13).
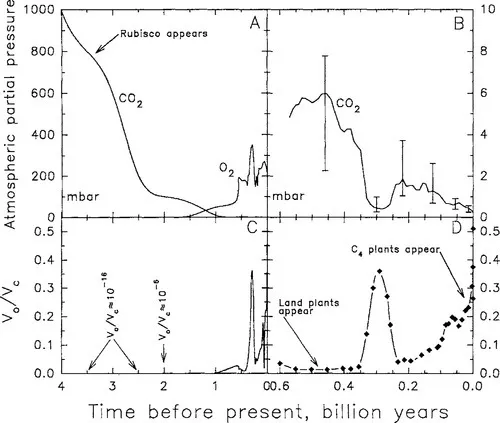
Figure 2 The modeled change in atmospheric CO2 and O2 partial pressures over (A) the past 4 billion years; and (B) the past 600 million years (according to Berner, 1994). C and D present the modeled change in Rubisco oxygenase activity (v0) to carboxylase activity (vc) at 30°C for the corresponding CO2 and O2 levels presented in A and B, respectively, calculated assuming a spinach C3-type Rubisco according to Jordan and Ogren, 1984. (adapted from Kasting, 1987, and Berner and Canfield, 1989)
Above 200 mbar oxygen, CO2 partial pressures of less than 500 μbar pose two problems. First, as a substrate for Rubisco carboxylation, CO2 availability becomes strongly limiting, reducing the turnover of the enzyme in vivo and imposing a limitation on photosynthesis by reducing the capacity of Rubisco to consume RuBP (Fig. 3A; Sage, 1995). Second, O2 competition becomes significant at warmer temperature (>30°C), and this causes a high rate of photorespiration (Fig. 3B; Sharkey, 1988; Sage, 1995). Whereas a CO2-substrate deficiency occurs only when the capacity for Rubisco to consume RuBP limits the rate of CO2 assimilation in plants, photorespiration is inhibitory regardless of whether Rubisco capacity or the capacity of the leaf to regenerate RuBP limits photosynthesis (Sharkey, 1985). At current CO2 levels, photorespiration can reduce photosynthesis by more than 40% at warmer temperatures (Sharkey, 1988; Ehleringer et al., 1991).
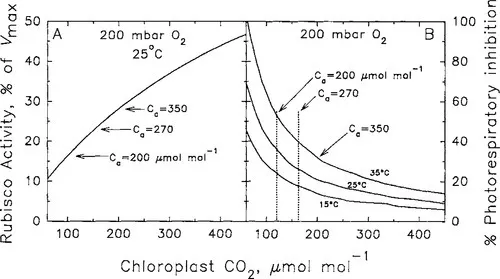
Figure 3 The modeled response of (A) Rubisco activity (as a percent of Vmax) as a function of stromal CO2 concentration, and (B) the percent of photorespiratory inhibition of photosynthesis (0.5v0/vc, × 100%). Modeled according to Sage (1995) using equations from Farquhar and von Caemmerer (1982). Ca indicates atmospheric CO2 content corresponding to indicated chloroplast CO2 concentrations. (Note: at sea level, μmol mol−1 ≅ μbar.)
The rise in photorespiratory potential triggered by the increase in the atmospheric O2:CO2 over the past 50 million years created high evolutionary pressure for dealing with the consequences of Rubisco oxygenation (Ehleringer et al., 1991). In all higher photosynthetic organisms, however, an elaborate biochemical edifice was already built around Rubisco, such that substantial barriers likely prevented the evolution of a novel carboxylase to replace Rubisco. Not only would the new carboxylase be required, but the accompanying biochemistry to regenerate acceptor molecules and process photosynthetic products would likely have to change as well. Such new photosynthetic systems would then have to compete against preexisting Rubisco-based systems, which though inefficient, would have had the advantage of working reasonably well and of being integrated into the associated cellular, organismal, and ecological systems. To a large degree, the potential for RuBP oxygenation represents a systematic constraint around which evolution must work, an evolutionary spandrel sensu Gould and Lewontin (1979) (Somerville et al., 1983). In a situation similar to that of a restoration architect who is constrained by preexisting structures, evolution is constrained by preexisting enzymes, genes, and regulatory systems in dealing with novel challenges (Gould and Lewontin, 1979). In plants and algae, the evolutionary response to declin...