
- 160 pages
- French
- ePUB (adaptée aux mobiles)
- Disponible sur iOS et Android
eBook - ePub
À propos de ce livre
L'évolution est-elle la transformation matérielle et graduelle des êtres vivants? Ou bien les espèces évoluent-elles en passant par des formes essentielles, presque des archétypes ? Pour connaître une espèce, est-ce le type qu'il faut caractériser ou la transformation qu'il faut expliquer ? L'un des meilleurs spécialistes au monde des dinosaures se fait ici historien pour nous apporter sa vision de ce problème clé de la théorie de l'évolution. Ce débat fournit à Kevin Padian l'occasion de revenir aux réflexions de Darwin sur la classification des espèces, mais aussi de nous faire redécouvrir un jalon important dans l'histoire de la pensée biologique, Sir Richard Owen, grand rival aujourd'hui oublié de l'auteur de L'Origine des espèces . Kevin Padian est professeur au département de biologie et au Muséum de paléontologie de l'Université de Californie, à Berkeley.
Approuvé par les 375,005 étudiants
Accès à plus d'un million de titres pour un prix mensuel raisonnable.
Étudiez plus efficacement en utilisant nos outils d'étude.
Informations
Sujet
Sciences biologiquesSujet
BiologieChapitre IV
Typologie et transformisme :
deux perspectives
sur les dinosaures et l’origine
des oiseaux
deux perspectives
sur les dinosaures et l’origine
des oiseaux
Lors de l’histoire de la reconstitution des animaux éteints, il y a une tension ou opposition intellectuelle entre ce que l’on peut appeler la typologie proprement dite et le transformisme. Il faut d’abord souligner à nouveau qu’il existe une différence considérable dans le sens du mot « typologie » en anglais et en français – c’est vraiment un « faux ami » pour les traducteurs. En français, la notion de typologie recouvre à peu près la description neutre d’une variabilité observée ou possible. Par exemple, on parle de la typologie microstructurale des tissus osseux – tels les tissus lamellaires avasculaires ou vasculaires, à ostéones primaires ou pas, ou les tissus fibro-lamellaires, dont les canaux vasculaires ont des orientations soit longitudinales, réticulaires, ou circulaires, etc. On emploie le terme pour décrire, différencier et classer la variation structurale naturelle.
En anglais, et surtout en américain, le terme « typologie » a une connotation tout à fait différente et moins neutre. Cherchons un peu dans l’histoire de la biologie évolutive, spécialement dans le cadre de la « théorie synthétique de l’évolution » à partir des années 1940. Nous constatons qu’une partie très importante en est le développement de ce qu’il est convenu d’appeler la « nouvelle systématique », surtout mise en avant par le professeur Ernst Mayr. Comme celui-ci l’a souvent souligné, l’important était d’établir un concept populationnel de l’espèce, par opposition à ce qu’il appelle le concept typologique de l’espèce. Par « populationnel », il voulait dire un concept « biologique » de l’espèce qui puisse refléter et valoriser les faits de variabilité naturelle des caractères au sein des populations, c’est-à-dire la véritable matière première de l’évolution. Il voulait donc se débarrasser d’un concept de l’espèce plus fixe, plus essentialiste, marqué par la typologie classique développée par Richard Owen, et dévalorisant la variabilité.
Mais alors que tout biologiste savait bien qu’il existe dans chaque espèce une variabilité naturelle, grâce à laquelle l’évolution elle-même peut avoir lieu, demeurait toujours le lancinant problème de l’association de la systématique avec des concepts plus ou moins fixes de l’espèce, concepts hérités de Linné, et cela ne serait-ce que pour des raisons pratiques de détermination et de nomenclature.
Par exemple, dans le cadre de la pratique taxinomique, il faut désigner un « échantillon type », pour pouvoir nommer une nouvelle espèce. Ce type ou porte-nom doit être conservé dans un musée public, et doit être accessible pour pouvoir être comparé à d’autres échantillons à déterminer. C’est là une question de pratique, de convenance, d’histoire, mais surtout de faits. Le problème est que chacun sait que cet échantillon type ne peut pas représenter toute la variation possible de l’espèce correspondante ni aujourd’hui, ni dans le passé, ni dans l’avenir. Ainsi, par analogie, l’image du « type » – qui était devenue assez platonicienne dans l’esprit d’Owen –, mais surtout par extension l’idée de typologie en général, est-elle devenue dangereuse, voire maudite, pour la « nouvelle systématique » et la « synthèse moderne » en ce que, du fait de son essentialisme, elle paraissait ignorer ou même contredire l’évolution.
De plus, lors de la fondation de la génétique classique, il y a une centaine d’années, régnait l’idée, surtout en Amérique et en Angleterre, qu’on ne devrait pas trouver beaucoup de variabilité génétique au sein des populations sauvages. Et ce parce que l’on croyait généralement que la plupart des mutations génétiques avaient des effets délétères ou létaux. Mayr, parmi d’autres, a insisté sur l’idée qu’il fallait au contraire beaucoup de variabilité génétique naturelle pour constituer la « matière première » de l’évolution dans les populations. Cette vision a été spécialement revendiquée pendant les années 1960, quand Thomas Jukes et Jack King découvrirent ce que l’on appela l’« évolution non darwinienne », c’est-à-dire le fait qu’il existe une énorme variabilité dans chaque génome spécifique et que la plupart des mutations correspondantes n’ont pas du tout d’effets graves – elles sont sensiblement neutres face à la sélection. C’est pour cela que l’on a parlé de « neutralisme » ou d’« évolution moléculaire neutre ». Ce genre de données a fourni, bien qu’un peu tardivement, une série d’arguments supplémentaires à l’encontre de la « typologie » proprement dite, arguments fondés cette fois sur la théorie génétique.
En résumé, la typologie conserve une connotation assez sulfureuse et négative dans la littérature scientifique en anglais, alors que ce terme peut avoir en français un sens beaucoup plus neutre et descriptif.
Je voudrais exposer ici certains travaux récents, aussi bien de ma part que de celle du professeur Jack Horner, de l’Université du Montana. Je vais parler un peu aussi, au passage, de notre travail en commun avec le professeur Armand de Ricqlès, qui est l’expert mondial sur la répartition et l’évolution de la micro-anatomie des tissus osseux des vertébrés fossiles. Je n’ai malheureusement qu’assez peu de temps pour vous expliquer l’ensemble de nos recherches communes, qui se poursuivent depuis presque une décennie. Toutefois j’espère que vous aurez l’occasion de suivre les œuvres du professeur de Ricqlès, qui plus que personne peut décrire et expliquer tout le sujet.
Je voudrais d’abord expliquer un peu le débat actuel à propos de la typologie – au sens anglais du terme, que je continuerai à employer – opposée au transformisme, tel qu’il est connu et discuté dans les pays anglophones. D’abord, je vous confie que c’est un débat qui demeure paradoxalement à peu près caché, y compris des participants eux-mêmes. Et il le demeure, je crois, à cause de préconceptions implicites conditionnées par la théorie. Comme je l’ai déjà raconté, selon les philosophes et les historiens des sciences, chacun demeure aveugle à ses propres préjugés théoriques. Ici nous allons rencontrer à ce propos un problème très intéressant, pouvant tout éclairer à la fois. Que pouvons-nous savoir – et comment – de la biologie d’animaux complètement éteints, sans aucun animal analogue actuel, sortes de chimères entre des types connus aujourd’hui, mais à la fois assez différents et originaux pour défier toute catégorisation ?
Considérons d’abord l’idée que, pour mieux étudier et reconstituer la biologie des dinosaures, il faut commencer par les traiter comme des reptiles actuels ordinaires, bien qu’immenses : de « bons reptiles », pour ainsi dire. Cela découle de l’idée très simple que, puisqu’on classe les dinosaures parmi les reptiles, ils ont dû posséder la plupart des caractéristiques typiquement reptiliennes. Donc, on peut commencer par postuler que leurs modes de croissance, leurs stratégies écologiques et leurs comportements étaient fondamentalement ceux des reptiles actuels, à quelque chose près. Nos collègues qui souscrivent à cette idée considèrent a priori qu’il ne faut pas même défendre ou soutenir ce point de vue par des données particulières, parce qu’il constitue en quelque sorte l’hypothèse par défaut. Toutefois, selon eux, il y a aussi beaucoup de données qui le soutiennent. On peut nommer ce point de vue « typologique » au sens anglais. Il se fonde a priori sur le fait que les dinosaures étant classés au sein des reptiles, il en découle nécessairement qu’ils devaient posséder des caractéristiques fonctionnelles, écologiques, ou physiologiques reptiliennes.
Considérons à présent l’antithèse qui pourrait être que les dinosaures étaient biologiquement moins comparables aux reptiles typiques d’aujourd’hui qu’aux oiseaux et ceux mammifères. Enfin, n’oublions pas que la diversité biologique de ce qui nous entoure aujourd’hui n’est qu’une ombre pâle de l’immense diversité du passé. Rappelons-nous que plus de 99,999 % de tous des organismes qui se sont succédé depuis le commencement du Phanérozoïque sont éteints aujourd’hui. Pourquoi présumer que notre diversité actuelle en est bien la représentation fidèle ? Et, dans ces conditions, comment reconstituer correctement les caractéristiques biologiques des organismes éteints ? Évidemment, si les organismes actuels ne constituent qu’une représentation aussi partielle de la biodiversité passée, c’est que l’extinction a éliminé la plupart des formes de transition, ce que Darwin avait bien compris. Dans ces conditions, pour pouvoir reconstituer la biologie des animaux éteints, il faut d’abord reconstituer leur généalogie, afin de mieux connaître la répartition systématique des caractéristiques que nous savons liées aux processus fonctionnels, écologiques et physiologiques.
Nous commencerons par la prémisse selon laquelle la méthode de construction de la classification nommée « phylogénétique » ou « cladistique » demeure le meilleur guide pour établir la parenté entre les êtres vivants. J’ai expliqué dans le premier chapitre que cette approche se fonde seulement sur la généalogie. On peut reconstituer celle-ci par l’analyse des caractéristiques évolutives nouvelles, ou synapomorphies, partagées par les codescendants étroitement apparentés d’un ancêtre commun exclusif. On peut retrouver ainsi l’ordre de descendance, indépendamment du degré de ressemblance global qui est beaucoup moins informatif. Comment se servir de la phylogénie pour mieux reconstituer les modes de vie des organismes éteints ?
Les hypothèses qui concernent les conditions physiologiques ou les comportements chez les organismes éteints ne peuvent être contrôlées ou testées directement que par deux moyens. Tout d’abord, il faudrait montrer que la présence d’une certaine structure (ou d’autres caractéristiques que l’on puisse examiner chez les fossiles) est à la fois nécessaire et suffisante pour que la condition physiologique correspondante soit réalisée. Deuxièmement, la présence de la structure susdite doit être aussi nécessaire et suffisante, et la fonction correspondante avérée chez un animal actuellement vivant – c’est ce que l’on nomme l’« hypothèse actualiste ». Jusqu’à présent, nous ne disposons d’aucun indicateur de ce type dans la documentation paléontologique pour décider de la physiologie chez un fossile. Tous les indicateurs disponibles sont indirects, et donc seulement plus ou moins indirectement liés avec un certain type de physiologie thermo-métabolique. En dépit de plusieurs affirmations publiées, aucun caractère anatomique particulier n’est jamais « corrélé causalement » ou ne constitue pas une « pierre de Rosette » pour indiquer directement et sans ambiguïté le métabolisme chez les vertébrés éteints. Ainsi, la présence ou l’absence d’un seul trait ou caractère lui-même plus ou moins lié à la physiologie a moins de poids qu’un faisceau de plusieurs traits indépendants mais concordants, surtout s’ils sont bien situés relativement les uns par rapport aux autres au sein d’un canevas phylogénétique établi indépendamment.
Les hypothèses concernant la manière par laquelle une condition donnée a évolué au long d’une lignée phylogénétique sont des hypothèses de processus. Habituellement, elles sont guidées et délimitées par ce que l’on croit savoir de la biologie des organismes vivants. On admet parfois que l’évolution peut étendre les limites des caractéristiques biologiques chez certaines formes éteintes si celles-ci paraissent, par certains caractères, situées au-delà de la distribution morphologique connue chez les animaux vivants, bien sûr. On peut construire des hypothèses sur l’évolution des telles conditions extrêmes, mais comment savoir si l’une est plus vraisemblable que d’autres ? Un contrôle efficace est d’examiner le patron évolutif de la lignée impliquée, à partir d’une phylogénie fondée sur des caractéristiques indépendantes. Les patrons phylogénétiques, strictement conçus, ne se basent pas sur des hypothèses concernant des processus évolutifs, ni sur l’appréciation de la plus ou moins grande plausibilité ou probabilité de certains d’entre eux. Ainsi la phylogénie elle-même devient un moyen de contrôle indépendant des hypothèses fondées sur les processus, bien qu’elle ne puisse jamais en devenir le juge ultime.
Par exemple, une hypothèse non enracinée dans la phylogénie pourrait proposer que les dinosaures auraient été ectothermes – c’est-à-dire plus ou moins à sang froid – simplement parce qu’ils étaient des reptiles et non pas des mammifères. En revanche, on peut construire un diagramme phylogénétique, ou cladogramme, autrement dit une sorte de plan, ou patron, de l’évolution de ce groupe, où l’on peut replacer diverses caractéristiques liées à la physiologie et au comportement, et tirer de cette structure logique les points successifs à partir desquels ces caractéristiques en question trouveraient leur origine. Pour se servir de cette approche, il faut que chaque trait soit partagé uniquement, et qu’il soit lié causalement et sans ambiguïté à des conséquences métaboliques. Ainsi on ne considère pas des caractéristiques isolées, mais la répartition phylogénétique globale de tout un écheveau de caractéristiques qui doivent se disposer de façon logique et additive sur le cladogramme entier.
Discutons un peu quelques approches plus ou moins typologiques de la question, étant entendu qu’il restera impossible de trouver une unanimité d’avis sur les questions qui regardent la biologie d’organismes fossiles, car l’observation directe en est impossible. Soyons conscients aussi que les observations elles-mêmes ne sont pas exemptes de préjugés du fait des convictions théoriques a priori, comme nous l’avons déjà vu. D’habitude, les discussions sur diverses questions de biologie des dinosaures en tant qu’organismes vivants commencent par des explications concernant l’approche philosophique des auteurs, ce qui nous intéresse ici presque autant que les données fournies et les conclusions elles-mêmes.
Paladino, Spotila et Dodson, des physiologistes et paléontologues, nous ont proposé une perspective raisonnée selon laquelle les dinosaures n’auraient été physiologiquement que des versions gigantesques des grands reptiles vivants tels le varan de Komodo et les tortues marines. Leur approche est d’étudier les processus métaboliques chez les reptiles actuels et d’en extrapoler les taux et valeurs jusqu’aux tailles des dinosaures. C’est tout à fait défendable, bien sûr ; c’est véritablement une voie pour la reconstitution biologique des formes éteintes – à condition qu’il s’agisse bien de formes exactement comparables aux organismes actuellement vivants. Mais comment le savoir ?
Pour commencer, Paladino et ses collègues suggèrent que quelques paléontologues se sont beaucoup trop appuyés sur l’approche phylogénétique dans leurs recherches paléophysiologiques. Puisque les dinosaures éteints sont phylogénétiquement plus proches des oiseaux que des crocodiles, ces paléontologues peuvent-ils à bon droit se laisser aller au préjugé, ou à la conclusion, que les oiseaux sont de meilleurs modèles physiologiques que ne le sont les crocodiles pour reconstituer les dinosaures ? Probablement à tort, disent-ils, parce que les représentants primitifs d’une lignée sont probablement plus semblables entre eux qu’ils ne le sont des formes plus avancées et spécialisées. (Nous serions d’accord, mais seulement si de telles ressemblances étaient fonctionnellement associées aux traits métaboliques spécialisés ou évolués, et non pas généraux ou primitifs de la lignée.) Remarquons que ces auteurs rejettent toute l’idée que l’analyse phylogénetique, qui exprime pourtant l’ordre généalogique des transitions de forme en forme au sein d’une lignée, puisse avoir une pertinence quelconque dans la résolution du problème.
Comme exemple de ce qu’ils considèrent comme les pièges de la méthode phylogénetique, Paladino et ses collègues discutent le célèbre et classique aphorisme cladistique selon lequel un dipneuste est plus voisin généalogiquement de la vache que de la truite (voir figure 3). Cette conclusion nous écarte de la physiologie, disent-ils, parce que le dipneuste et la truite partagent beaucoup plus de traits primitifs, ce qui confirme l’ancien système linnéen, qui les regarde tous deux comme des poissons, contrairement à la vache. Mais peut-être n’est-ce pas un bon exemple. Tout d’abord, il ne nous faut pas un cladogramme pour montrer que les vaches diffèrent assez par leur physiologie à la fois du dipneuste et de la truite ; cela ne nous avance pas beaucoup. Deuxièmement, c’est un fait que les dipneustes partagent quelques caractéristiques physiologiquement importantes avec tous les tétrapodes – et pas seulement les vaches (qui constituent un exemple très dérivé de tétrapodes, soulignons-le). Par exemple, les dipneustes et tous les tétrapodes partagent des choanes internes, des poumons fonctionnels, des membres charnus avec un seul os à la base, et la capacité de survivre à l’assèchement saisonnier de leur milieu. Nous ne savons pas si la physiologie des premiers tétrapodes ressemblait davantage à celle des dipneustes actuels qu’à celles des tétrapodes actuels, mais ils pouvaient réaliser des fonctions physiologiques que ne peuvent jamais réaliser les truites, ni anciennes ni actuelles. Et par ces traits nouveaux, insolites, hérités de leur ancêtre commun, les sarcoptérygiens ont pu réaliser puis perfectionner les mécanismes physiologiques permettant la survie hors de l’eau et la locomotion terrestre. C’est pour cela qu’on propose de se servir de la phylogénie, et non de l’ignorer.
Paladino et ses collègues remarquent que les oiseaux actuels n’ont pas de dents et ont perdu la longue queue osseuse des reptiles (bien qu’ils admettent que « certains oiseaux mésozoïques » les avaient encore), et donc ces caractères constituent « deux autres traits par lesquels les dinosaures ressemblent plus aux crocodiliens qu’aux oiseaux modernes ». Ainsi, on devrait en conclure que les dinosaures étaient fonctionnellement plus proches des crocodiles que des oiseaux. Mais, si les oiseaux primitifs retenaient une longue queue osseuse et des dents, pourquoi n’étaient-ils pas eux-mêmes plus comparables aux crocodiles qu’aux oiseaux actuels – même s’ils possédaient des plumes (et nous savons maintenant que les dinosaures les plus proches des oiseaux en possédaient aussi) ? Ce manque de logique souligne le danger d’abandonner la perspective phylogénétique en cherchant à résoudre des questions évolutives.
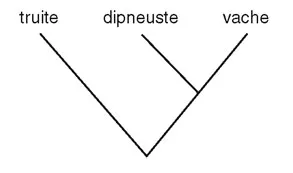
Figure 3
Malgré ces limitations, Paladino et ses collègues produisent une abondance d’informations comparatives sur la physiologie des animaux actuels, et ils les extrapolent pour les dinosaures éteints, en se fondant sur les principes élémentaires de l’uniformitarisme et sur les données concernant les formes éteintes. Ils conclurent que les dinosaures pouvaient très bien fonctionner simplement comme des versions gigantesques des reptiles ectothermes connus aujourd’...
Table des matières
- Couverture
- Titre
- Copyright
- Avertissement
- Préface - par Armand de Ricqlès
- Chapitre I - Darwin et la systématique : aux origines de la pensée phylogénétique
- Chapitre II - Richard Owen : ambiguïté et richesse des transformismes prédarwiniens
- Chapitre III - Typologie ou phylogénie : de l’invention du concept de dinosaure à son évolution actuelle
- Chapitre IV - Typologie et transformisme : deux perspectives sur les dinosaures et l’origine des oiseaux
- Références bibliographiques
- Dans la même collection
Foire aux questions
Oui, vous pouvez résilier à tout moment à partir de l'onglet Abonnement dans les paramètres de votre compte sur le site Web de Perlego. Votre abonnement restera actif jusqu'à la fin de votre période de facturation actuelle. Découvrir comment résilier votre abonnement
Non, les livres ne peuvent pas être téléchargés sous forme de fichiers externes, tels que des PDF, pour être utilisés en dehors de Perlego. Cependant, vous pouvez télécharger des livres dans l'application Perlego pour les lire hors ligne sur votre téléphone portable ou votre tablette. Apprendre à télécharger des livres hors ligne
Perlego propose deux abonnements : Essentiel et Complet
- Essentiel est idéal pour les étudiants et les professionnels qui aiment explorer un large éventail de sujets. Accédez à la bibliothèque Essentiel comprenant plus de 800 000 titres de référence et best-sellers dans les domaines du commerce, du développement personnel et des sciences humaines. Il comprend un temps de lecture illimité et la voix standard de la fonction Écouter.
- Complet est parfait pour les étudiants avancés et les chercheurs qui ont besoin d'un accès complet et illimité. Accédez à plus de 1,4 million de livres sur des centaines de sujets, y compris des titres académiques et spécialisés. L'abonnement Complet comprend également des fonctionnalités avancées telles que la fonction Écouter Premium et l'Assistant de recherche.
Nous sommes un service d'abonnement à des ouvrages universitaires en ligne, où vous pouvez accéder à toute une bibliothèque pour un prix inférieur à celui d'un seul livre par mois. Avec plus d'un million de livres sur plus de 990 sujets, nous avons ce qu'il vous faut ! En savoir plus sur notre mission
Recherchez le symbole Écouter sur votre prochain livre pour voir si vous pouvez l'écouter. L'outil Écouter lit le texte à haute voix pour vous, en surlignant le passage qui est en cours de lecture. Vous pouvez le mettre sur pause, l'accélérer ou le ralentir. En savoir plus sur la fonctionnalité Écouter
Oui ! Vous pouvez utiliser l'application Perlego sur les appareils iOS et Android pour lire à tout moment, n'importe où, même hors ligne. Parfait pour les trajets quotidiens ou lorsque vous êtes en déplacement.
Veuillez noter que nous ne pouvons pas prendre en charge les appareils fonctionnant sur iOS 13 et Android 7 ou versions antérieures. En savoir plus sur l'utilisation de l'application
Veuillez noter que nous ne pouvons pas prendre en charge les appareils fonctionnant sur iOS 13 et Android 7 ou versions antérieures. En savoir plus sur l'utilisation de l'application
Oui, vous pouvez accéder à De Darwin aux dinosaures par Kevin Padian en format PDF et/ou ePUB ainsi qu'à d'autres livres populaires dans Sciences biologiques et Biologie. Nous disposons de plus d'un million d'ouvrages à découvrir dans notre catalogue.