
- 240 pages
- French
- ePUB (adaptée aux mobiles)
- Disponible sur iOS et Android
eBook - ePub
Les Neurones miroirs
À propos de ce livre
À l'évidence, faire quelque chose et imaginer le faire ne reviennent pas au même. Et pourtant ! Il se pourrait bien que, pour notre cerveau, la pensée et l'action soient une seule et même chose. Voilà ce que révèlent Giacomo Rizzolatti et son équipe, qui ont découvert des neurones étonnants : ils s'activent lorsqu'on effectue une action, mais aussi lorsqu'on voit quelqu'un d'autre la réaliser lui-même. Ce livre décrit les stupéfiantes propriétés de ces « neurones miroirs », explique leur mécanisme et souligne leur importance. Ils sont les promoteurs du langage, ils expliquent pourquoi nous parlons aussi avec nos mains. Ils rendent compte de l'expression des émotions ; ils sont le mécanisme de notre compréhension d'autrui. Au terme de ce parcours inédit dans le cerveau, une interrogation surgit : et si ces neurones miroirs étaient à la base de nos comportements sociaux ?Giacomo Rizzolatti, professeur de physiologie humaine, dirige le département de neurosciences de l'Université de Parme. Corrado Sinigaglia est professeur de philosophie des sciences à l'Université de Milan.
Approuvé par les 375,005 étudiants
Accès à plus de 1,5 million de titres pour un prix mensuel raisonnable.
Étudiez plus efficacement en utilisant nos outils d'étude.
Informations
Sujet
Biological SciencesSujet
NeuroscienceChapitre 1
Le système moteur
Prendre une tasse à café
Commençons par un exemple. Rien ne paraît plus simple que prendre en main une tasse à café. Pourtant, ce geste si naturel comporte une multiplicité de processus tellement enchevêtrés qu’à première vue ils nous semblent indistincts. Tout d’abord, nous devons identifier la tasse en la choisissant parmi les autres objets éventuels qui nous entourent et qui se disputent notre attention. Pour ce faire, nous devons tourner la tête et les yeux de telle sorte que l’image de la tasse tombe exactement sur la fovéa, la zone où l’acuité visuelle est maximale, nous permettant ainsi d’en apprécier au mieux les différents aspects (sa forme, l’orientation de son anse, sa couleur, etc.). Si nous voulons la prendre, nous devons ensuite la localiser par rapport à notre corps : ce n’est qu’alors que nous pourrons allonger la main et l’atteindre. Dans le même temps, nous devons en prendre, pour ainsi dire, les mesures, afin de pouvoir la saisir de la façon que nous considérons comme la plus opportune.
La tasse nous dicte un ensemble de mesures et de modalités de préhension : c’est à nous de répondre et de décider comment bouger et nous y conformer, en choisissant parmi toutes les prises possibles la plus adaptée à l’utilisation que nous voulons en faire ou éventuellement la plus conforme à nos habitudes. Bien que, d’ordinaire, nous ne nous en apercevions pas, avant même de l’atteindre, les doigts et la paume de notre main ont déjà commencé à se représenter la configuration géométrique de la portion de la tasse qui nous intéresse et les éventuels types de préhension qui lui sont corrélés. Dès que nous l’atteignons, notre main reçoit des informations venant de la peau, des articulations et des muscles qui lui permettent d’affiner sa prise et de porter la tasse à la bouche.
Même sans considérer les ajustements de posture – qui anticipent la réalisation de chacun des mouvements décrits ci-dessus, en en prévoyant les conséquences et en contrôlant l’équilibre dynamique du corps dans les différentes situations où il agit –, et indépendamment du rôle joué par l’apprentissage et l’expérience dans les différentes phases d’identification, de localisation, de déplacement de la main et de préhension de l’objet, un geste aussi élémentaire que saisir une tasse à café révèle un entrelacement complexe de sensations (visuelles, tactiles, olfactives, proprioceptives, etc.), de motivations, de dispositions corporelles et de performances motrices qui interagissent entre elles et avec les objets qui nous entourent, en réalisant chaque fois des formes de syntonies plus ou moins subtiles.
Mais que se passe-t-il lorsque nous passons du plan descriptif au plan neurophysiologique ? Devons-nous nous attendre à ce que les différents processus énumérés ci-dessus soient réductibles à des circuits corticaux distincts du point de vue anatomique et fonctionnel ? Dès lors, quels systèmes neuronaux sont concernés au niveau cortical lorsque nous nous décidons à prendre une tasse à café, et comment ces systèmes coopèrent-ils entre eux ?
L’organisation des aires motrices frontales
Il se peut que le lecteur soit surpris par la simplicité de notre exemple, et du fait qu’il ressortit aux neurosciences, voire aux sciences cognitives. Pourtant, ces vingt dernières années, l’analyse des mécanismes neuronaux qui sous-tendent un acte aussi élémentaire que la préhension (ou que les nombreux autres actes de notre vie quotidienne) nous a conduits à reconsidérer bien des aspects essentiels de notre façon traditionnelle de concevoir le fonctionnement du cerveau – en particulier, concernant l’organisation du système moteur et les relations fonctionnelles qu’il entretient avec les autres systèmes (sensoriels, mais pas seulement), où se déploie l’activité cérébrale.
On a longtemps considéré que les phénomènes sensoriels, perceptifs et moteurs étaient répartis dans des aires corticales nettement distinctes : d’un côté, les aires sensorielles, à savoir les aires visuelles (localisées dans le lobe occipital), somato-sensorielles (cortex somato-sensoriel), auditives (circonvolutions temporales supérieures), etc. ; de l’autre, les aires motrices, situées dans la partie postérieure du lobe frontal, connue également sous le nom de cortex frontal agranulaire. Entre les premières et les deuxièmes se situent de vastes régions corticales, souvent définies comme aires associatives : à celles-ci (en particulier aux aires temporo-pariétales) serait impartie la tâche de « rassembler » les informations qui proviennent des différentes aires sensorielles et de former des « percepts » d’objets spatiaux qui doivent être transmis aux aires motrices pour l’organisation des différents mouvements (figure 1.1).
Selon ce modèle, lorsque nous prenons un objet avec la main, notre cerveau effectuerait un ensemble de processus organisés de façon sérielle, au cours desquels les informations provenant des aires corticales postérieures (sensorielles) seraient intégrées par les aires associatives ; le résultat de l’élaboration de ces dernières serait ensuite transmis au cortex moteur pour la réalisation de mouvements appropriés, dont l’exécution effective dépendrait de l’intention explicite du sujet.
Le système moteur aurait donc un rôle périphérique et éminemment exécutif – comme le montrent, du reste, les cartographies fonctionnelles qui figurent dans la plupart des manuels de neurologie encore en vogue aujourd’hui. Ainsi, par exemple, il suffit de penser au classique simiunculus de Clinton Woolsey, ou au non moins célèbre homunculus de Wilder Penfield (figure 1.2), datant de la moitié du XXe siècle, obtenus par une stimulation électrique du cortex moteur à l’aide de macro-électrodes disposées à sa surface1. Tant C. Woolsey que W. Penfield distinguaient deux aires motrices : l’aire motrice primaire (AMP) et l’aire motrice supplémentaire (AMS), caractérisées par une représentation complète des mouvements, plus détaillée dans l’aire motrice primaire, plus grossière dans l’aire motrice supplémentaire.Toutefois, les cartographies obtenues par une stimulation électrique de surface ne correspondaient pas totalement à l’organisation cytoarchitectonique de la partie postérieure du lobe frontal (cortex moteur) des primates qui avait été décrite au début du XXe siècle par Korbinian Brodmann2. Il divisait cette région du lobe frontal en deux aires distinctes (aire 4 et aire 6), sur la base de la distribution des cellules pyramidales de la couche V (figure 1.3). L’aire AMP incluait, en effet, la totalité de l’aire 4 et une grande partie de l’aire 6, placée sur la face latérale de l’hémisphère, tandis que l’aire AMS coïncidait avec la portion de l’aire 6 située sur la face mésiale. Pour remédier à cette difficulté, Clinton Woolsey suggéra que la différence cytoarchitectonique entre l’aire 4 et l’aire 6 ne correspondait à aucune distinction fonctionnelle, mais uniquement à une représentation somato-topique différente. Selon Clinton Woolsey, les mouvements de la main, les mouvements fins de la bouche et du pied (mouvements distaux) étaient localisés dans l’aire 4, tandis que les mouvements des bras, des jambes (mouvements proximaux) et du tronc (axiaux) le seraient dans l’aire 6.
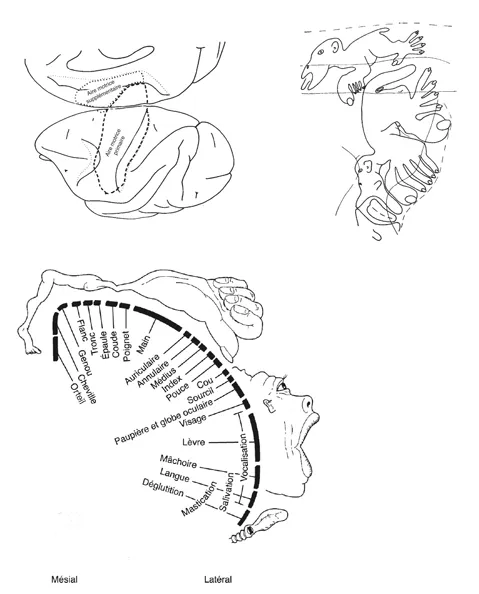
Fig. 1.2. En haut à gauche : vision mésiale et latérale du cerveau du singe ; les lignes en tirets indiquent la localisation de l’aire motrice primaire et de l’aire motrice supplémentaire selon la neurologie classique. En haut, à droite : les deux demi-homoncules correspondant, respectivement, à la représentation somato-topique des mouvements dans le cortex moteur primaire et dans le cortex moteur supplémentaire, selon la description de Clinton Woolsey. En bas, à gauche : l’homoncule moteur de Wilder Penfield.
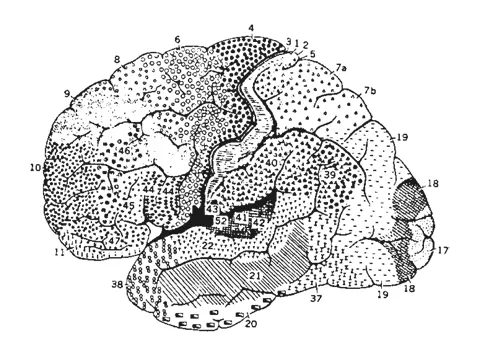
Fig. 1.3. Cartes cytoarchitectoniques du cortex d’un singe (cercopithèque, à gauche) et d’un homme (à droite). Ces cartes ont été obtenues par K. Brodmann grâce à des méthodes histologiques qui permettaient de colorier sélectivement les éléments cellulaires et, donc, de distinguer les différentes aires du cortex cérébral sur la base du nombre de couches corticales (typiquement six), de leur grandeur, de la quantité de neurones contenus et de la distribution de trois types fondamentaux de neurones corticaux (cellules pyramidales, étoilées et fusiformes). Si nous comparons la carte corticale du singe à celle de l’homme, nous pouvons remarquer qu’il existe une similitude de fond. Les mêmes sillons principaux (sillon central, sillon latéral, sillon temporel supérieur) sont présents dans les deux cartes et, à quelques exceptions près, les mêmes aires cytoarchitectoniques. Cependant, il existe aussi des différences importantes. Par exemple, la région pariéto-temporo-occipitale est beaucoup plus grande chez l’homme que chez le singe, ce qui a induit un déplacement des aires visuelles : dans le cortex de l’homme, elles occupent la face mésiale de l’hémisphère, tandis que, dans celui du singe, elles occupent une grande partie du pôle caudal des hémisphères cérébraux sur la surface latérale (aire 17). En outre, le cortex humain révèle un développement important du lobe frontal. En revanche, il existe chez le singe un sillon qui ne se retrouve pas chez l’homme : le sillon arqué. Il divise le lobe frontal en deux portions cytoarchitectoniquement et fonctionnellement différentes. La portion postérieure, formée par les aires 4 et 6, est caractérisée par une absence quasi totale de cellules étoilées (dites également « granules ») et par l’absence de la quatrième couche (couche granulaire interne) d’où son nom de cortex agranulaire. Bien que l’examen macroscopique ne révèle pas chez l’homme une subdivision du lobe frontal si marquée, des études cytoarchitectoniques et des analyses fonctionnelles ont montré que, dans ce cas aussi, il est possible de distinguer une partie postérieure et une partie antérieure, la première (aires 4 et 6) est dévolue principalement à l’activité motrice, la seconde (souvent désignée comme lobe préfrontal) a des fonctions de type cognitif.
Malgré les nombreuses critiques de ce qui fut jugé par certains comme une solution ad hoc, le schéma général des deux simi-homoncules constitua pendant des années un des points incontournables de la neurologie. Et cela pour au moins deux raisons : tout d’abord, ce schéma fournissait une explication immédiate et facilement applicable sur le plan clinique des problèmes de la localisation des mouvements dans le cortex moteur ; en outre, il reflétait l’idée, aujourd’hui comme alors très répandue, de l’unité fonctionnelle du système moteur cortical, véritable point d’arrivée de l’information sensorielle élaborée par les aires associatives et qui, en soi, serait privé de toute compétence perceptive et cognitive.
Pour reprendre les termes d’Elwood Henneman, ce système au fond n’existerait dans le cerveau que « pour traduire des pensées et des sensations en mouvement3 ». Mais comment et où se produit une telle traduction ? Autrement dit, à quel moment la pensée et la perception cessent-elles d’être telles pour se transformer en mouvement ? Et Elwood Henneman d’ajouter : « Pour l’heure, les premiers pas de ce processus demeurent hors de portée de notre analyse4. » Toutefois, à peine quelques années plus tard (1984), on a commencé à comprendre que le système moteur n’est pas seulement connecté anatomiquement aux aires corticales responsables des activités cérébrales impliquées dans les « pensées et les sensations », mais qu’il possède de multiples fonctions, lesquelles ne sauraient être enfermées dans le cadre d’une cartographie purement exécutive.
Loin d’être organisé uniquement en deux aires (AMP et AMS), le cortex moteur est formé d’une constellation de régions différentes5. Si nous comparons la parcellisation anatomico-fonctionelle actuelle du cortex agranulaire, telle qu’elle est représentée dans la figure 1.4 avec les cartographies proposées dans les figures 1.2 et 1.3, nous pouvons remarquer que, contrairement aux hypothèses de Clinton Woolsey, le cortex moteur primaire (AMP, à partir de maintenant F1), coïncide avec l’aire 4 de Brodmann. Quant à l’aire 6, elle apparaît divisée en trois régions principales (mésiale, dorsale, ventrale), lesquelles sont à leur tour subdivisées en une partie rostrale (antérieure) en une partie caudale (postérieure) : la région mésiale est formée par les aires F3 (AMS) et F6 (pré-AMS) ; la région dorsale (cortex prémoteur dorsal) par les aires F2 (P...
Table des matières
- Couverture
- Titre
- Introduction
- Chapitre 1 - Le système moteur
- Chapitre 2 - Le cerveau qui agit
- Chapitre 3 - L’espace autour de nous
- Chapitre 4 - Agir et comprendre
- Chapitre 5 - Les neurones miroirs chez l’homme
- Chapitre 6 - Imitation et langage
- Chapitre 7 - Le partage des émotions
- Bibliographie
- Remerciements
- Cahier photo
- Index
Foire aux questions
Oui, vous pouvez résilier à tout moment à partir de l'onglet Abonnement dans les paramètres de votre compte sur le site Web de Perlego. Votre abonnement restera actif jusqu'à la fin de votre période de facturation actuelle. Découvrir comment résilier votre abonnement
Non, les livres ne peuvent pas être téléchargés sous forme de fichiers externes, tels que des PDF, pour être utilisés en dehors de Perlego. Cependant, vous pouvez télécharger des livres dans l'application Perlego pour les lire hors ligne sur votre téléphone portable ou votre tablette. Apprendre à télécharger des livres hors ligne
Perlego propose deux formules : Essential et Complete
- Essential est idéal pour les apprenants et les professionnels qui aiment explorer une grande variété de sujets. Accédez à la Essential Library avec plus de 800 000 titres de confiance et best-sellers dans les domaines du business, du développement personnel et des sciences humaines. Inclut un temps de lecture illimité et la voix Standard Read Aloud.
- Complete : Parfait pour les apprenants avancés et les chercheurs ayant besoin d'un accès total et sans restriction. Débloquez plus de 1,5 million de livres dans des centaines de sujets, y compris des titres académiques et spécialisés. Le forfait Complete inclut aussi des fonctionnalités avancées telles que Premium Read Aloud et Research Assistant.
Nous sommes un service déabonnement à des manuels scolaires en ligne, qui vous permet d'accéder à une bibliothèque en ligne entière pour moins que le prix d'un seul livre par mois. Avec plus de 1,5 million de livres sur plus de 990 thèmes, nous avons ce qu'il vous faut ! Découvrir notre mission
Recherchez le symbole Écouter sur votre prochain livre pour voir si vous pouvez l'écouter. L'outil Écouter lit le texte à haute voix pour vous, en surlignant le passage qui est en cours de lecture. Vous pouvez le mettre sur pause, l'accélérer ou le ralentir. En savoir plus sur la fonctionnalité Écouter
Oui ! Vous pouvez utiliser l'application Perlego sur les appareils iOS et Android pour lire à tout moment, n'importe où, même hors ligne. Parfait pour les trajets quotidiens ou lorsque vous êtes en déplacement.
Veuillez noter que nous ne pouvons pas prendre en charge les appareils fonctionnant sur iOS 13 et Android 7 ou versions antérieures. En savoir plus sur l'utilisation de l'application
Veuillez noter que nous ne pouvons pas prendre en charge les appareils fonctionnant sur iOS 13 et Android 7 ou versions antérieures. En savoir plus sur l'utilisation de l'application
Oui, vous pouvez accéder à Les Neurones miroirs de Giacomo Rizzolatti,Corrado Sinigaglia aux formats PDF et/ou ePub, ainsi qu'à d'autres livres populaires dans Biological Sciences et Neuroscience. Nous avons plus de 1,5 million de livres disponibles dans notre catalogue pour vous.