The new and fully updated edition of the definitive haematology textbook for undergraduate and postgraduate students and trainees
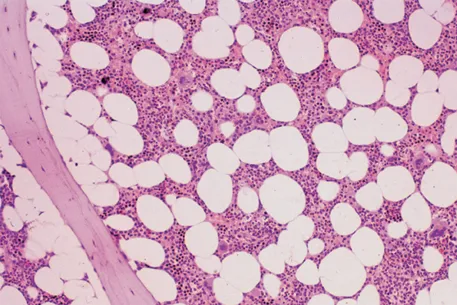
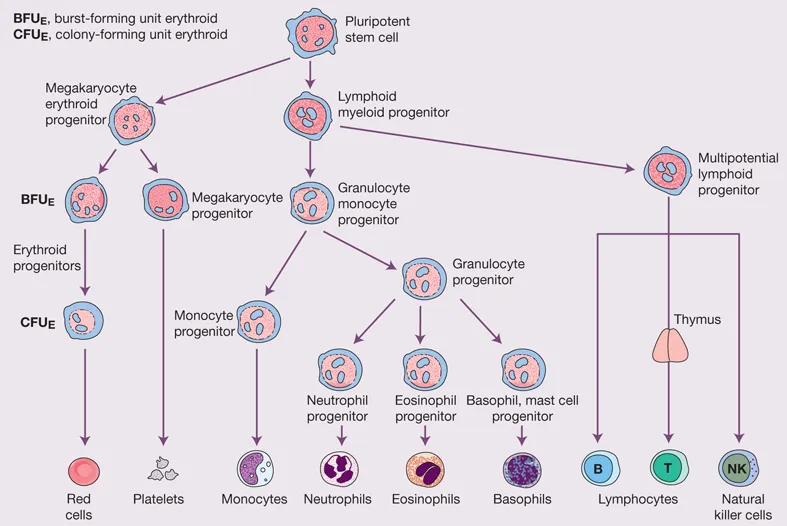
Hoffbrand's Essential Haematology is widely regarded as the most authoritative introduction to the subject available, helping medical students and trainee doctors understand the essential principles of modern clinical and laboratory haematology for nearly four decades. Now in its eighth edition, this market-leading textbook introduces the formation and function of blood cells and the diseases that arise from dysfunction and disruption of these processes.
Beautifully presented with over 300 stunning colour illustrations, the new edition has been thoroughly updated to reflect recent advances in knowledge of the pathogenesis of blood diseases and their diagnosis and treatment. This new text:
- Describes disorders and diseases of the blood such as the various anaemias and white cell disorders, leukaemias, lymphomas and myeloma, as well as bleeding and thrombotic disorders
- Incorporates the latest World Health Organization (WHO) classification of haematological neoplastic diseases
- Reviews contemporary application of multiparameter flow cytometry, DNA sequencing and other technologies in evaluating patients with suspected haematological disease
- Discusses the therapeutic use of chimeric antigen T-cells, mono- and bi-specific monoclonal antibodies, inhibitors of intracellular signalling pathways and direct orally acting anticoagulants
- Includes sections on blood transfusion and the haematological aspects of systemic diseases, pregnancy and the neonate
Hoffbrand's Essential Haematology is a vital resource for all students and trainees, and a valuable reference for practicing specialists wishing to update their knowledge.