
- 600 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Molecular Biology of B Cells
About this book
Molecular Biology of B Cells, Second Edition is a comprehensive reference to how B cells are generated, selected, activated and engaged in antibody production. All of these developmental and stimulatory processes are described in molecular, immunological, and genetic terms to give a clear understanding of complex phenotypes.
Molecular Biology of B Cells, Second Edition offers an integrated view of all aspects of B cells to produce a normal immune response as a constant, and the molecular basis of numerous diseases due to B cell abnormality. The new edition continues its success with updated research on microRNAs in B cell development and immunity, new developments in understanding lymphoma biology, and therapeutic targeting of B cells for clinical application. With updated research and continued comprehensive coverage of all aspects of B cell biology, Molecular Biology of B Cells, Second Edition is the definitive resource, vital for researchers across molecular biology, immunology and genetics.
- Covers signaling mechanisms regulating B cell differentiation
- Provides information on the development of therapeutics using monoclonal antibodies and clinical application of Ab
- Contains studies on B cell tumors from various stages of B lymphocytes
- Offers an integrated view of all aspects of B cells to produce a normal immune response
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 1
The Structure and Regulation of the Immunoglobulin Loci
Joseph S. Lucas1, Cornelis Murre1, Ann J. Feeney2, and Roy Riblet3 1Division of Biological Sciences, Department of Molecular Biology, University of California, San Diego, CA, USA 2Department of Immunology and Microbial Science, The Scripps Research Institute, CA, USA 3Torrey Pines Institute for Molecular Studies, San Diego, CA, USA
Abstract
The temporal and lineage specificity of antigen receptor assembly is regulated at multiple levels. These include locus conformation, germline transcription, chromatin remodeling, epigenetic marking, and nuclear location. Here we discuss how the immunoglobulin loci are structured and how the assembly of antigen receptor elements during B cell development is regulated.
Keywords
3D-structure; Antibody repertoire; B cell development; Combinatorial diversity; Epigenetic marking; Germline transcription; Immunoglobulin; Junctional diversity; Transcription factor; V(D)J recombination1. Introduction
Our adaptive immune system relies heavily on the use of antibodies, produced by B cells, to eliminate foreign pathogens and toxins. It is estimated that mammalian organisms have the ability to generate on the order of 1011 different antibodies [1]. This staggering number is made possible by an elaborate mechanism that assembles complete antigen binding site sequences from arrays of gene segments encoding portions of the complete antibody proteins. This assembly involves combinatorial selection of each type of gene segment and creation of short novel sequences at the junctions of these DNA segments, thus generating a vast variety of assembled gene sequences and antigen binding specificities. Sequence diversity is further increased by the somatic hypermutation process that is active in these sequences.
Antibodies are proteins made up of two identical heavy chains and two identical light chains. All heavy chains are expressed from the Igh locus, whereas light chains are expressed from one of two loci, Igκ or Igλ [2]. The DNA segments that rearrange to create antibody heavy and light chain genes in developing B cells include variable (V) and joining (J) elements, and heavy chain genes include a third, diversity (Dh), gene segment [3]. Single light chain V and J and heavy chain V, D, and J segments are selected and joined in a seemingly random process out of the many such genomic segments that span as much as 3 Mbp of DNA
Over the past 30 years, a large number of studies have been performed to describe the process of V(D)J recombination in molecular terms, including how it is regulated during development; how it is controlled by cell signaling; how it is modulated by transcription; and how chromatin modifications, nuclear positioning and three-dimensional (3D) topology affect rearrangement. Collectively, these experiments aimed to address a number of critical questions. Some of the prominent questions follow: How do DNA sequences separated by millions of base pairs interact to allow recombination? How are gene segments selected randomly to allow for the generation of antibody diversity?
2. Genomic Organization of the Mouse Immunoglobulin Heavy Chain Locus
The Igh locus is made up of multiple variable (Vh), Dh, joining (Jh), and constant (Ch) segments, arrayed in adjacent regions of the Igh locus. In mice the locus spans 2.75 million base pairs, and there are about 100 Vh segments that have seemingly functional coding sequences and nearly as many nonfunctional Vh segments, 10–15 Dh segments depending on mouse strain, and four Jh segments. Eight constant regions encode the Igh isotypes [4,5]. Vh genes encode most of the heavy chain variable region including the first two hypervariable loops that form the binding site for antigen [6,7]. Dh segments are very short, 10–15 nucleotides of coding sequence, but they are critically important in the generation of antibody diversity. Dh segments determine most of the heavy chain’s third hypervariable region or complementarity determining region 3 (CDR3) [8–10]. In the folded protein’s antigen binding site, this region makes major, sometimes dominant, contacts with antigen.
Each set of gene segments has evolved, and continues to evolve, via gene addition by duplication, divergence through mutation, and loss by nonsense or frameshift mutations or by partial or total deletion. A prominent example is the ancestral duplication of the constant region IgG2a gene and divergence and alternate gene deletion leaving the present IgG2a gene in BALB/c and many other mouse strains and the IgG2c gene in C57BL/6 and related strains [11,12]. A larger scale case in the Igh-V (Vh) array is the duplication of the entire proximal region of the Vh array comprising the interspersed Vh7183 and VhQ52 genes (Ighv5 and Ighv2 families in Figure 1) from 400 kb in C57BL/6 to 800 kb in 129 and BALB/c [5].
Alignment of Vh gene sequences and grouping them by similarity yields an evolutionary structure of the gene population, revealing three major branches or groups, splitting further into 15 families of genes that share 80% sequence identity. The three groups of Vh genes are an ancient evolutionary division; Vh genes from any vertebrate species fall into several or all three of these groups. Vh gene family structure is apparently not generally shared beyond closely related species [13], although the Vh7183 or Ighv5 family is found widely [14]. In mice, at least, a family of Vh genes occupies a restricted region of the Vh array and are interspersed there with other families; the proximal, 3′ 300 kb region of the Vh locus contains only the interspersed Vh7183 and VhQ52 (Ighv5 and Ighv2) families, the distal, 5′ 1.5 Mb portion contains only VhJ558 and Vh3609 (Ighv1 and Ighv8) genes, and the central 680 kb contains the remaining families.
In developing B cells, Vh, Dh, and Jh segments recombine to encode the antigen-binding domain of the antibody heavy chain [3]. Each Vh, Dh, and Jh segment is flanked by a short DNA sequence called the recombination signal sequence (RSS). This sequence is recognized by recombination activating gene enzymes RAG1 and RAG2. RAG1 and RAG2 form a complex with additional proteins and act to create a loop between the two RSSs, bringing them into proximity. The RAG complex then induces double-strand DNA breaks and promotes ligation of appropriate coding segments via the nonhomologous end-joining machinery [15–17].
In the Igh locus, V, D, J, and C gene segments are in separate clusters, and all in the same 5′→3′ polarity. VDJ rearrangements proceed via serial excisions of intervening sequences, D to J, then V to DJ, and loss of the excised sequences as free closed DNA circles. VDJ recombination is irreversible, although a secondary V–V replacement mechanism occurs at low frequency [18].
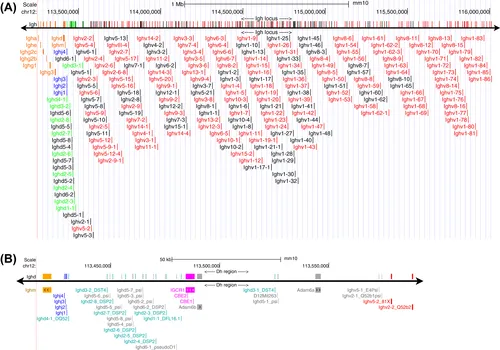
FIGURE 1 The genomic structure of the immunoglobulin heavy chain locus.
The 2.75 Mb Igh locus is located on the distal region of mouse chromosome 12. (A) The various types of gene segments are represented in different colors: functional Vh segments are red, functional Dh segments are green, the four Jh segments are blue, and the eight Ch loci are orange. Nonfunctional Vh and Dh pseudogenes are black. The sequence is from mm10, the Dec. 2011 Mus musculus assembly (Genome Reference Consortium Mouse Build 38), documented in NCBI Gene ID: 111507 [http://ncbi.nlm.nih.gov/gene/?term=111507]. (B) Expanded view of the Dh region; the IgM constant region at the left is orange; the Jh regions are blue; functional Dh segments are cyan; nonfunctional Dh, Vh, and Adam6 pseudogenes are gray; the intergenic control region 1 (88) with its two CCCTC-binding factor binding elements, CBE1 and CBE2, are magenta; and functional Vh genes at the right are red.
3. Genomic Organization of the Mouse Immunoglobulin Kappa Light Chain Locus
The kappa light chain locus (Figure 2) is even larger than the heavy chain locus, extending 3.17 Mb; it too contains approximately 100 functional and 60 nonfunctional V segments and four functional (plus one nonfunctional) J segments [19]. Light chain genes do not use D segments. Interestingly, half of all Vk genes are in the opposite polarity to the Jk and Ck loci and must rearrange through a nondestructive inversion of the portion of the Vk gene array between the joining Vk and Jk segments. As a result, many Vk segments are retained rather than excised, for potential use in a secondary rearrangement with any remaining downstream Jk segments. This conserves a larger set of Vk genes for “B cell editing,” i.e., replacement of an initial Vk-Jk choice that yields an antiself antibody or is otherwise a poor partner for the rearranged heavy chain. There seems to be no bias between the excision and inversion mechanisms because both sets of Vk genes contain similar frequencies of both functional genes and highly...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Dedication
- Preface
- Contributors
- Chapter 1. The Structure and Regulation of the Immunoglobulin Loci
- Chapter 2. The Mechanism of V(D)J Recombination
- Chapter 3. Transcriptional Regulation of Early B Cell Development
- Chapter 4. Relationships among B Cell Populations Revealed by Global Gene Analysis
- Chapter 5. Roles of MicroRNAs in B Lymphocyte Physiology and Oncogenesis
- Chapter 6. Proliferation and Differentiation Programs of Developing B Cells
- Chapter 7. Development and Function of B Cell Subsets
- Chapter 8. B Cells and Antibodies in Jawless Vertebrates
- Chapter 9. The Origin of V(D)J Diversification
- Chapter 10. Structure and Signaling Function of the B-Cell Antigen Receptor and Its Coreceptors
- Chapter 11. Fc and Complement Receptors
- Chapter 12. B Cell Localization and Migration in Health and Disease
- Chapter 13. B Cells as Regulators
- Chapter 14. B Cell Memory and Plasma Cell Development
- Chapter 15. The Role of the BAFF and Lymphotoxin Pathways in B Cell Biology
- Chapter 16. The Mucosal Immune System: Host–Bacteria Interaction and Regulation of Immunoglobulin A Synthesis
- Chapter 17. Gut Microbiota and Their Regulation
- Chapter 18. Molecular Mechanisms of AID Function
- Chapter 19. The Mechanism of IgH Class Switch Recombination
- Chapter 20. Somatic Hypermutation: The Molecular Mechanisms Underlying the Production of Effective High-Affinity Antibodies
- Chapter 21. Aberrant AID Expression by Pathogen Infection
- Chapter 22. Molecular Pathogenesis of B Cell Lymphomas
- Chapter 23. B Cells Producing Pathogenic Autoantibodies
- Chapter 24. The Cellular and Molecular Biology of HIV-1 Broadly Neutralizing Antibodies
- Chapter 25. Immune Deficiencies Caused by B Cell Defects
- Chapter 26. IMGT® Immunoglobulin Repertoire Analysis and Antibody Humanization
- Chapter 27. Anti-Interleukin-6 Receptor Antibody Therapy Against Autoimmune Inflammatory Diseases
- Chapter 28. Targeting the IL-17/IL-23 Axis in Chronic Inflammatory Immune-Mediated Diseases
- Chapter 29. Discovery and Development of Anti-TNF Therapy: Pillar of a Therapeutic Revolution
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Molecular Biology of B Cells by Tasuku Honjo,Michael Reth,Andreas Radbruch,Frederick Alt in PDF and/or ePUB format, as well as other popular books in Medicine & Immunology. We have over 1.5 million books available in our catalogue for you to explore.