
eBook - ePub
Molecular Biology of B Cells
Tasuku Honjo,Michael Reth,Andreas Radbruch,Frederick Alt
This is a test
Buch teilen
- 600 Seiten
- English
- ePUB (handyfreundlich)
- Über iOS und Android verfügbar
eBook - ePub
Molecular Biology of B Cells
Tasuku Honjo,Michael Reth,Andreas Radbruch,Frederick Alt
Angaben zum Buch
Buchvorschau
Inhaltsverzeichnis
Quellenangaben
Über dieses Buch
Molecular Biology of B Cells, Second Edition is a comprehensive reference to how B cells are generated, selected, activated and engaged in antibody production.All of these developmental and stimulatory processes are described in molecular, immunological, and genetic terms to give a clear understanding of complex phenotypes.
Molecular Biology of B Cells, Second Edition offers an integrated view of all aspects of B cells to produce a normal immune response as a constant, and the molecular basis of numerous diseases due to B cell abnormality. The new edition continues its success with updated research on microRNAs in B cell development and immunity, new developments in understanding lymphoma biology, and therapeutic targeting of B cells for clinical application. With updated research and continued comprehensive coverage of all aspects of B cell biology, Molecular Biology of B Cells, Second Edition is the definitive resource, vital for researchers across molecular biology, immunology and genetics.
- Covers signaling mechanisms regulating B cell differentiation
- Provides information on the development of therapeutics using monoclonal antibodies and clinical application of Ab
- Contains studies on B cell tumors from various stages of B lymphocytes
- Offers an integrated view of all aspects of B cells to produce a normal immune response
Häufig gestellte Fragen
Wie kann ich mein Abo kündigen?
Gehe einfach zum Kontobereich in den Einstellungen und klicke auf „Abo kündigen“ – ganz einfach. Nachdem du gekündigt hast, bleibt deine Mitgliedschaft für den verbleibenden Abozeitraum, den du bereits bezahlt hast, aktiv. Mehr Informationen hier.
(Wie) Kann ich Bücher herunterladen?
Derzeit stehen all unsere auf Mobilgeräte reagierenden ePub-Bücher zum Download über die App zur Verfügung. Die meisten unserer PDFs stehen ebenfalls zum Download bereit; wir arbeiten daran, auch die übrigen PDFs zum Download anzubieten, bei denen dies aktuell noch nicht möglich ist. Weitere Informationen hier.
Welcher Unterschied besteht bei den Preisen zwischen den Aboplänen?
Mit beiden Aboplänen erhältst du vollen Zugang zur Bibliothek und allen Funktionen von Perlego. Die einzigen Unterschiede bestehen im Preis und dem Abozeitraum: Mit dem Jahresabo sparst du auf 12 Monate gerechnet im Vergleich zum Monatsabo rund 30 %.
Was ist Perlego?
Wir sind ein Online-Abodienst für Lehrbücher, bei dem du für weniger als den Preis eines einzelnen Buches pro Monat Zugang zu einer ganzen Online-Bibliothek erhältst. Mit über 1 Million Büchern zu über 1.000 verschiedenen Themen haben wir bestimmt alles, was du brauchst! Weitere Informationen hier.
Unterstützt Perlego Text-zu-Sprache?
Achte auf das Symbol zum Vorlesen in deinem nächsten Buch, um zu sehen, ob du es dir auch anhören kannst. Bei diesem Tool wird dir Text laut vorgelesen, wobei der Text beim Vorlesen auch grafisch hervorgehoben wird. Du kannst das Vorlesen jederzeit anhalten, beschleunigen und verlangsamen. Weitere Informationen hier.
Ist Molecular Biology of B Cells als Online-PDF/ePub verfügbar?
Ja, du hast Zugang zu Molecular Biology of B Cells von Tasuku Honjo,Michael Reth,Andreas Radbruch,Frederick Alt im PDF- und/oder ePub-Format sowie zu anderen beliebten Büchern aus Medicina & Immunologia. Aus unserem Katalog stehen dir über 1 Million Bücher zur Verfügung.
Information
Chapter 1
The Structure and Regulation of the Immunoglobulin Loci
Joseph S. Lucas1, Cornelis Murre1, Ann J. Feeney2, and Roy Riblet3 1Division of Biological Sciences, Department of Molecular Biology, University of California, San Diego, CA, USA 2Department of Immunology and Microbial Science, The Scripps Research Institute, CA, USA 3Torrey Pines Institute for Molecular Studies, San Diego, CA, USA
Abstract
The temporal and lineage specificity of antigen receptor assembly is regulated at multiple levels. These include locus conformation, germline transcription, chromatin remodeling, epigenetic marking, and nuclear location. Here we discuss how the immunoglobulin loci are structured and how the assembly of antigen receptor elements during B cell development is regulated.
Keywords
3D-structure; Antibody repertoire; B cell development; Combinatorial diversity; Epigenetic marking; Germline transcription; Immunoglobulin; Junctional diversity; Transcription factor; V(D)J recombination1. Introduction
Our adaptive immune system relies heavily on the use of antibodies, produced by B cells, to eliminate foreign pathogens and toxins. It is estimated that mammalian organisms have the ability to generate on the order of 1011 different antibodies [1]. This staggering number is made possible by an elaborate mechanism that assembles complete antigen binding site sequences from arrays of gene segments encoding portions of the complete antibody proteins. This assembly involves combinatorial selection of each type of gene segment and creation of short novel sequences at the junctions of these DNA segments, thus generating a vast variety of assembled gene sequences and antigen binding specificities. Sequence diversity is further increased by the somatic hypermutation process that is active in these sequences.
Antibodies are proteins made up of two identical heavy chains and two identical light chains. All heavy chains are expressed from the Igh locus, whereas light chains are expressed from one of two loci, Igκ or Igλ [2]. The DNA segments that rearrange to create antibody heavy and light chain genes in developing B cells include variable (V) and joining (J) elements, and heavy chain genes include a third, diversity (Dh), gene segment [3]. Single light chain V and J and heavy chain V, D, and J segments are selected and joined in a seemingly random process out of the many such genomic segments that span as much as 3 Mbp of DNA
Over the past 30 years, a large number of studies have been performed to describe the process of V(D)J recombination in molecular terms, including how it is regulated during development; how it is controlled by cell signaling; how it is modulated by transcription; and how chromatin modifications, nuclear positioning and three-dimensional (3D) topology affect rearrangement. Collectively, these experiments aimed to address a number of critical questions. Some of the prominent questions follow: How do DNA sequences separated by millions of base pairs interact to allow recombination? How are gene segments selected randomly to allow for the generation of antibody diversity?
2. Genomic Organization of the Mouse Immunoglobulin Heavy Chain Locus
The Igh locus is made up of multiple variable (Vh), Dh, joining (Jh), and constant (Ch) segments, arrayed in adjacent regions of the Igh locus. In mice the locus spans 2.75 million base pairs, and there are about 100 Vh segments that have seemingly functional coding sequences and nearly as many nonfunctional Vh segments, 10–15 Dh segments depending on mouse strain, and four Jh segments. Eight constant regions encode the Igh isotypes [4,5]. Vh genes encode most of the heavy chain variable region including the first two hypervariable loops that form the binding site for antigen [6,7]. Dh segments are very short, 10–15 nucleotides of coding sequence, but they are critically important in the generation of antibody diversity. Dh segments determine most of the heavy chain’s third hypervariable region or complementarity determining region 3 (CDR3) [8–10]. In the folded protein’s antigen binding site, this region makes major, sometimes dominant, contacts with antigen.
Each set of gene segments has evolved, and continues to evolve, via gene addition by duplication, divergence through mutation, and loss by nonsense or frameshift mutations or by partial or total deletion. A prominent example is the ancestral duplication of the constant region IgG2a gene and divergence and alternate gene deletion leaving the present IgG2a gene in BALB/c and many other mouse strains and the IgG2c gene in C57BL/6 and related strains [11,12]. A larger scale case in the Igh-V (Vh) array is the duplication of the entire proximal region of the Vh array comprising the interspersed Vh7183 and VhQ52 genes (Ighv5 and Ighv2 families in Figure 1) from 400 kb in C57BL/6 to 800 kb in 129 and BALB/c [5].
Alignment of Vh gene sequences and grouping them by similarity yields an evolutionary structure of the gene population, revealing three major branches or groups, splitting further into 15 families of genes that share 80% sequence identity. The three groups of Vh genes are an ancient evolutionary division; Vh genes from any vertebrate species fall into several or all three of these groups. Vh gene family structure is apparently not generally shared beyond closely related species [13], although the Vh7183 or Ighv5 family is found widely [14]. In mice, at least, a family of Vh genes occupies a restricted region of the Vh array and are interspersed there with other families; the proximal, 3′ 300 kb region of the Vh locus contains only the interspersed Vh7183 and VhQ52 (Ighv5 and Ighv2) families, the distal, 5′ 1.5 Mb portion contains only VhJ558 and Vh3609 (Ighv1 and Ighv8) genes, and the central 680 kb contains the remaining families.
In developing B cells, Vh, Dh, and Jh segments recombine to encode the antigen-binding domain of the antibody heavy chain [3]. Each Vh, Dh, and Jh segment is flanked by a short DNA sequence called the recombination signal sequence (RSS). This sequence is recognized by recombination activating gene enzymes RAG1 and RAG2. RAG1 and RAG2 form a complex with additional proteins and act to create a loop between the two RSSs, bringing them into proximity. The RAG complex then induces double-strand DNA breaks and promotes ligation of appropriate coding segments via the nonhomologous end-joining machinery [15–17].
In the Igh locus, V, D, J, and C gene segments are in separate clusters, and all in the same 5′→3′ polarity. VDJ rearrangements proceed via serial excisions of intervening sequences, D to J, then V to DJ, and loss of the excised sequences as free closed DNA circles. VDJ recombination is irreversible, although a secondary V–V replacement mechanism occurs at low frequency [18].
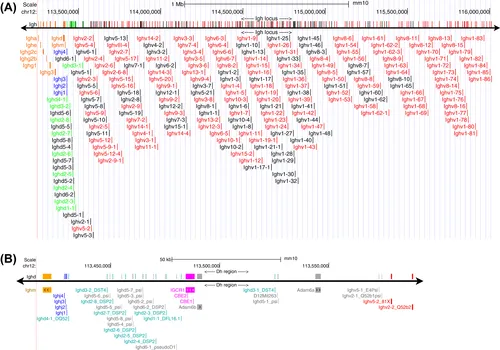
FIGURE 1 The genomic structure of the immunoglobulin heavy chain locus.
The 2.75 Mb Igh locus is located on the distal region of mouse chromosome 12. (A) The various types of gene segments are represented in different colors: functional Vh segments are red, functional Dh segments are green, the four Jh segments are blue, and the eight Ch loci are orange. Nonfunctional Vh and Dh pseudogenes are black. The sequence is from mm10, the Dec. 2011 Mus musculus assembly (Genome Reference Consortium Mouse Build 38), documented in NCBI Gene ID: 111507 [http://ncbi.nlm.nih.gov/gene/?term=111507]. (B) Expanded view of the Dh region; the IgM constant region at the left is orange; the Jh regions are blue; functional Dh segments are cyan; nonfunctional Dh, Vh, and Adam6 pseudogenes are gray; the intergenic control region 1 (88) with its two CCCTC-binding factor binding elements, CBE1 and CBE2, are magenta; and functional Vh genes at the right are red.
3. Genomic Organization of the Mouse Immunoglobulin Kappa Light Chain Locus
The kappa light chain locus (Figure 2) is even larger than the heavy chain locus, extending 3.17 Mb; it too contains approximately 100 functional and 60 nonfunctional V segments and four functional (plus one nonfunctional) J segments [19]. Light chain genes do not use D segments. Interestingly, half of all Vk genes are in the opposite polarity to the Jk and Ck loci and must rearrange through a nondestructive inversion of the portion of the Vk gene array between the joining Vk and Jk segments. As a result, many Vk segments are retained rather than excised, for potential use in a secondary rearrangement with any remaining downstream Jk segments. This conserves a larger set of Vk genes for “B cell editing,” i.e., replacement of an initial Vk-Jk choice that yields an antiself antibody or is otherwise a poor partner for the rearranged heavy chain. There seems to be no bias between the excision and inversion mechanisms because both sets of Vk genes contain similar frequencies of both functional genes and highly...