The new and fully updated edition of the definitive haematology textbook for undergraduate and postgraduate students and trainees
Hoffbrand's Essential Haematology is widely regarded as the most authoritative introduction to the subject available, helping medical students and trainee doctors understand the essential principles of modern clinical and laboratory haematology for nearly four decades. Now in its eighth edition, this market-leading textbook introduces the formation and function of blood cells and the diseases that arise from dysfunction and disruption of these processes.
Beautifully presented with over 300 stunning colour illustrations, the new edition has been thoroughly updated to reflect recent advances in knowledge of the pathogenesis of blood diseases and their diagnosis and treatment. This new text:
Describes disorders and diseases of the blood such as the various anaemias and white cell disorders, leukaemias, lymphomas and myeloma, as well as bleeding and thrombotic disorders
Incorporates the latest World Health Organization (WHO) classification of haematological neoplastic diseases
Reviews contemporary application of multiparameter flow cytometry, DNA sequencing and other technologies in evaluating patients with suspected haematological disease
Discusses the therapeutic use of chimeric antigen T-cells, mono- and bi-specific monoclonal antibodies, inhibitors of intracellular signalling pathways and direct orally acting anticoagulants
Includes sections on blood transfusion and the haematological aspects of systemic diseases, pregnancy and the neonate
Hoffbrand's Essential Haematology is a vital resource for all students and trainees, and a valuable reference for practicing specialists wishing to update their knowledge.
Foire aux questions
Comment puis-je résilier mon abonnement ?
Il vous suffit de vous rendre dans la section compte dans paramètres et de cliquer sur « Résilier l’abonnement ». C’est aussi simple que cela ! Une fois que vous aurez résilié votre abonnement, il restera actif pour le reste de la période pour laquelle vous avez payé. Découvrez-en plus ici.
Puis-je / comment puis-je télécharger des livres ?
Pour le moment, tous nos livres en format ePub adaptés aux mobiles peuvent être téléchargés via l’application. La plupart de nos PDF sont également disponibles en téléchargement et les autres seront téléchargeables très prochainement. Découvrez-en plus ici.
Quelle est la différence entre les formules tarifaires ?
Les deux abonnements vous donnent un accès complet à la bibliothèque et à toutes les fonctionnalités de Perlego. Les seules différences sont les tarifs ainsi que la période d’abonnement : avec l’abonnement annuel, vous économiserez environ 30 % par rapport à 12 mois d’abonnement mensuel.
Qu’est-ce que Perlego ?
Nous sommes un service d’abonnement à des ouvrages universitaires en ligne, où vous pouvez accéder à toute une bibliothèque pour un prix inférieur à celui d’un seul livre par mois. Avec plus d’un million de livres sur plus de 1 000 sujets, nous avons ce qu’il vous faut ! Découvrez-en plus ici.
Prenez-vous en charge la synthèse vocale ?
Recherchez le symbole Écouter sur votre prochain livre pour voir si vous pouvez l’écouter. L’outil Écouter lit le texte à haute voix pour vous, en surlignant le passage qui est en cours de lecture. Vous pouvez le mettre sur pause, l’accélérer ou le ralentir. Découvrez-en plus ici.
Est-ce que Hoffbrand's Essential Haematology est un PDF/ePUB en ligne ?
Oui, vous pouvez accéder à Hoffbrand's Essential Haematology par Victor Hoffbrand, David P. Steensma en format PDF et/ou ePUB ainsi qu’à d’autres livres populaires dans Médecine et Hématologie. Nous disposons de plus d’un million d’ouvrages à découvrir dans notre catalogue.
This first chapter is concerned with the general aspects of blood cell formation (haemopoiesis). The processes that regulate haemopoiesis and the early stages of formation of red cells (erythropoiesis), granulocytes and monocytes (myelopoiesis) and platelets (thrombopoiesis) are also discussed.
Site of haemopoiesis
In the first few weeks of gestation, the embryonic yolk sac is a transient site of haemopoiesis called ‘primitive haemopoiesis’. However, ‘definitive haemopoiesis’ derives from a population of stem cells first observed on the aorta-gonads-mesonephros (AGM) region of the developing embryo. These common precursors of endothelial and haemopoietic cells are called haemangioblasts and are believed to seed the liver, spleen and bone marrow.
From 6 weeks until 6–7 months of fetal life, the liver and spleen are the major haemopoietic organs and continue to produce blood cells until about 2 weeks after birth (Table 1.1; see Fig. 7.1b). The placenta also contributes to fetal haemopoiesis. The bone marrow is the most important site from 6–7 months of fetal life. During normal childhood and adult life, the marrow is the only source of new blood cells. The developing cells are situated outside the bone marrow sinuses; mature cells are released into the sinus spaces, the marrow microcirculation and so into the general circulation.
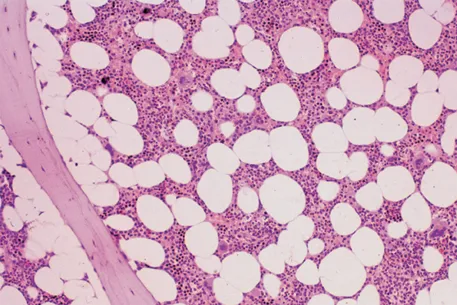
In infancy all the bone marrow is haemopoietic, but during childhood and beyond there is progressive fatty replacement of marrow throughout the long bones, so that in adult life haemopoietic marrow is confined to the central skeleton and proximal ends of the femurs and humeri (Table 1.1). Even in these active haemopoietic areas, approximately 50% of the marrow consists of fat in the middle-aged adult (Fig.1.1). The remaining fatty marrow is capable of reversion to haemopoiesis and in many diseases there is also expansion of haemopoiesis down the long bones. Moreover, in certain disease states the liver and spleen can resume their fetal haemopoietic role (‘extramedullary haemopoiesis’).
Table 1.1Dominant sites of haemopoiesis at different stages of development.
Fetus
0–2 months (yolk sac)
2–7 months (liver, spleen)
5–9 months (bone marrow)
Infants
Bone marrow (practically all bones); dwindling post-parturition contribution from liver/spleen that ceases in the first few months of life
Adults
Vertebrae, ribs, sternum, skull, sacrum and pelvis, proximal ends of femur
Figure 1.1 Normal bone marrow trephine biopsy (posterior iliac crest). Haematoxylin and eosin stain; approximately 50% of the intertrabecular tissue is haemopoietic tissue and 50% is fat.
Haemopoietic stem and progenitor cells
Haemopoiesis starts with a pluripotential stem cell that can self-renew by asymmetrical cell division, but also gives rise to the separate cell lineages. These cells are able to repopulate a bone marrow from which all stem cells have been eliminated by lethal irradiation or chemotherapy. Self-renewal and repopulating ability define the haemopoietic stem cell (HSC). HSCs are rare, perhaps 1 in every 20 million nucleated cells in bone marrow. Newer DNA sequencing techniques suggest that a typical adult has approximately 50 000 HSCs.
HSCs are heterogeneous, with some able to repopulate a bone marrow for more than 16 weeks, called long-term HSCs, while others, although able to produce all haemopoietic cell types, engraft only transiently for a few weeks and are called short–term HSCs. Although the exact cell surface marker phenotype of the HSC is still unknown, on immunological testing these cells are positive for the marker Cluster of Differentiation 34 (CD34+) and negative for CD38− and for cell lineage-defining markers (Lin−). Morphologically, HSCs have the appearance of a small or medium-sized lymphocyte (see Fig. 23.3). The cells reside adjacent to osteoblasts or to endothelial cells of sinusoidal vessels in endosteal or vascular ‘niches’, where they are surrounded by stromal cells, with which they interact in numerous ways. The niches also contain sympathetic nerve endings.
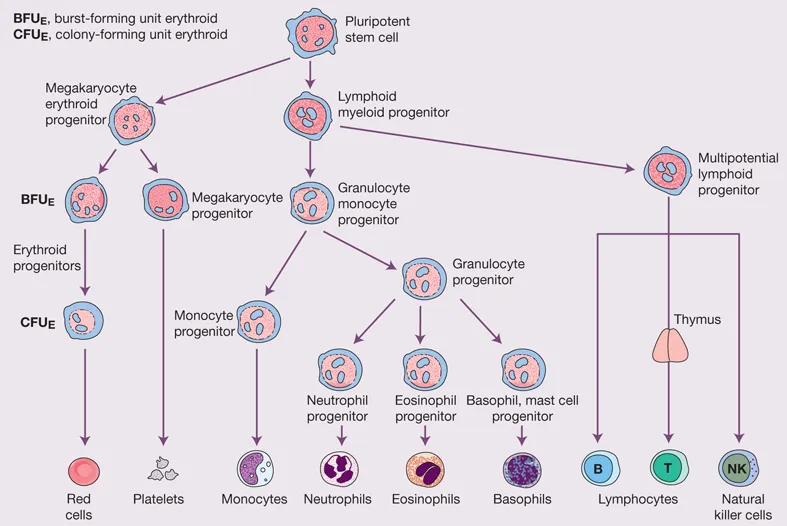
Cell differentiation occurs from the stem cells via committed haemopoietic progenitors, which are restricted in their developmental potential (Fig. 1.2). The existence of the separate progenitor cells can be demonstrated by in vitro culture techniques. Stem cells and very early progenitors are assayed by culture on bone marrow stroma as long-term culture- initiating cells, whereas late progenitors are generally assayed in semi-solid media. As examples, in the erythroid series progenitors can be identified in special cultures as burst-forming units (BFU-E, describing the ‘burst’ with which they form in culture) and colony-forming units (CFU-E; Fig 1.2); the mixed granulocyte/monocyte progenitor is identified as a colony-forming unit-granulocyte/monocyte (CFU-GM) in culture. Megakaryocytes form from the CFU-Meg.
Figure 1.2 Diagrammatic representation of the bone marrow pluripotent stem cells (haemopoietic stem cells, HSC) and the cell lines that arise from them. A megakaryocytic/erythroid progenitor (MkEP) and a mixed lymphoid/myeloid progenitor are formed from the pluripotent stem cells. Each gives rise to more differentiated progenitors. The MkEP divides into erythroid and megakaryocyte progenitors. The mixed lymphoid progenitor gives rise to B and T lymphocytes and to natural killer cells. A granulocyte/monocyte progenitor gives rise to progenitors for monocytes, neutrophils, eosinophils, basophils and mast cells. The erythroid progenitors are also termed BFU-E and CFU-E. BFU-E, burst-forming unit erythroid; CFU-E, colony-forming unit erythroid.
In the haemopoietic hierarchy, the pluripotent stem cell gives rise to a mixed erythroid and megakaryocyte progenitor, which then divides into separate erythroid and megakaryocyte progenitors. The pluripotent stem cell also gives rise to a mixed lymphoid, granulocyte and monocyte progenitor, which divides into a progenitor of granulocytes and monocytes and a mixed lymphoid progenitor, from which B- and T-cell lymphocytes and natural killer (NK) cells develop (Fig. 1.2). The spleen, lymph nodes and thymus are secondary sites of lymphocyte production (see Chapter 9).
The stem cell has the capability for self-renewal (Fig. 1.3), so that...