![]()
1. L’apparato muscolare scheletrico
Il tessuto muscolare scheletrico è costituito da fibrocellule che, funzionalmente, si suddividono in:
-
Fibre rapide
-
Fibre intermedie
-
Fibre lente
I muscoli che sviluppano grande potenza e velocità sono fondamentalmente composti da fibre rapide. Le caratteristiche delle fibre rapide sono:
-
grandezza maggiore,
-
reticolo sarcoplasmatico più sviluppato (rilascio più rapido di calcio),
-
elevata quantità di enzimi glicolitici,
-
minor densità capillare (metabolismo ossidativo ridotto),
-
minor numero di mitocondri per il medesimo motivo.
Di contro quelle lente sono:
-
più piccole,
-
innervate da fibre nervose più piccole,
-
più vascolarizzate,
-
ad alto numero di mitocondri,
-
ad alto contenuto di mioglobina.
E’ proprio la mioglobina (la cui cinetica di cessione dell’ossigeno è ben diversa da quella dell’emoglobina) che conferisce il colore rossastro a queste fibre, alle quali è stato dato il nome di fibre rosse mentre quelle rapide sono dette bianche.
E’ quindi evidente che le fibre rosse sono adatte alle prestazioni atletiche di durata o al sostegno del corpo contro la gravità (variazioni di tono), mentre le bianche saranno utilizzate per le attività esplosive di scatto e salto e per le contrazioni muscolari rapide e veloci.
Secondo alcuni non risulta che l’allenamento possa cambiare la proporzione relativa tra fibre lente e veloci: sarebbero quindi fondamentalmente i fattori genetici a dire se un soggetto è più o meno adatto a certe specialità. Nondimeno, agobiopsie sul quadricipite di atleti di alto livello hanno messo in luce queste proporzioni limite:
A partire dai 45 anni l’allenamento permette di mantenere in vita fibre veloci, che tendono a diminuire, trasformandole in fibre lente.
Basi molecolari della contrazione
Le fibre muscolari scheletriche e cardiache presentano la caratteristica striatura, dovuta alla giustapposizione dei sarcomeri, tenuti insieme da proteine strutturali (Strie Z). L’unità funzionale del muscolo scheletrico e cardiaco è il sarcomero. Esso è costituito da filamenti sottili, ancorati alle strie Z e protrudenti verso il centro del sarcomero e da filamenti spessi, a cavallo della linea mediana (Figura). Questi ultimi sono costituiti da fasci di molecole di miosina, unite coda a coda da altre proteine strutturali della linea mediana, e sono rivolte verso il rispettivo emisarcomero. I filamenti sottili sono costituiti da tre proteine: actina, tropomiosina e troponina. Queste due sono dette proteine modulanti o regolatorie e dalla loro situazione sterica dipende la possibilità che l’actina reagisca con le teste della miosina, a formare un ponte trasverso, un legame transiente fra le due (acto-miosina) che permette sia lo scivolamento dei filamenti sottili sugli spessi (con variazione di lunghezza), sia lo sviluppo di forza.
Dal muscolo ai miofilamenti
Perché ciò avvenga sono necessari due “attivatori” della contrazione:
-
lo ione calcio (Ca2+) è l’attivatore del filamento sottile, in quanto, legandosi con la subunità C della troponina, permette la disinibizione del sito attivo dell’actina, fino a quel momento mascherato dalla subunità I, disponibile così a legarsi alla testa della miosina;
-
l’ADP, prodotto dall’idrolisi dell’ATP dall’attività della testa della miosina, e l’energia che ne deriva, sono gli attivatori del filamento spesso.
Il Ca2+ entrato attraverso la membrana cellulare grazie al potenziale d’azione stimola la fuoriuscita di Ca2+ dal reticolo sarcoplasmatico e questo fenomeno è responsabile della rapida attivazione delle unità contrattili. L’ATP di per sé ha effetto di rilasciamento, si sganciano cioè i ponti appena formati e solo dopo la demolizione di una nuova molecola ad ADP + Pi (fosfato inorganico) + E (energia) si potranno formare altri ponti. L’ATP è dunque responsabile dell’effetto di rigor.
La teoria consolidata e comunemente accettata della contrazione muscolare è quella cosiddetta dello scorrimento dei miofilamenti (Sir Andrew Huxley). I ponti trasversi si formano a una determinata velocità, che sarà poi proporzionale alla velocità di contrazione del muscolo in toto, che dipende, a sua volta, dalla velocità di idrolisi dell’ATP. Di conseguenza le fibre veloci possiederanno un’attività enzimatica delle teste della miosina (ATPasi miosinica) mediamente più veloce che quella delle fibre lente. In effetti la miosina, come tante proteine, è rappresentata in più isoforme, dalle lente alle veloci, e sono queste che condizioneranno la velocità di contrazione e quindi la potenza, in quanto Forza x Velocità = Potenza.
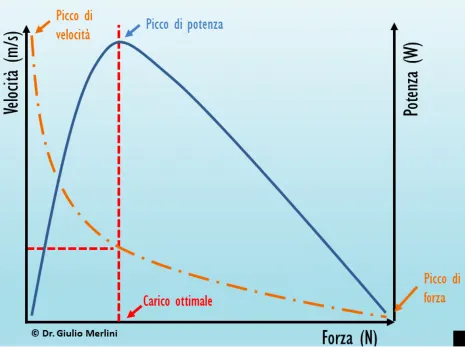
Il rapporto fra forza, velocità e potenza di contrazione muscolare è ben rappresentato nel diagramma Forza/Velocità.
Figura da https//www.corebosport.com: Relazione Forza/Velocità.
Il diagramma ci dice che la velocità è massima quanto più basso è il carico, il che significa passare da una velocità massima (vmax), virtualmente a carico zero, ma, in realtà un carico esiste sempre ed è dato dalla quota-peso dell’arto o segmento corporeo mossi dal muscolo, a una velocità zero, quando la forza sviluppata è massima (curva spezzata). La massima forza di un muscolo si sviluppa in contrazione isometrica (v. infra), dove la velocità di accorciamento esterno del muscolo è proprio nulla. Moltiplicando punto a punto i valori di forza e velocità si ottiene il profilo della potenza (curva blu), che sarà vicina a zero quando forza o velocità sono vicine a zero e raggiungerà il suo massimo quando la forza è all’incirca al 30% di Fmax. Questo è un dato da tener presente quando si desidera allenare e potenziare un muscolo. In Fisiologia muscolare la forza (F) viene più spesso riportata come tensione (T), che è la forza riferita alla sezione trasversa del muscolo e permette di confrontare la prestazione di muscoli di tutte le dimensioni. Questo ci porta al diagramma Tensione/Lunghezza, che rappresenta il meccanismo cardine della contrattilità scheletrica e cardiaca (Figura).
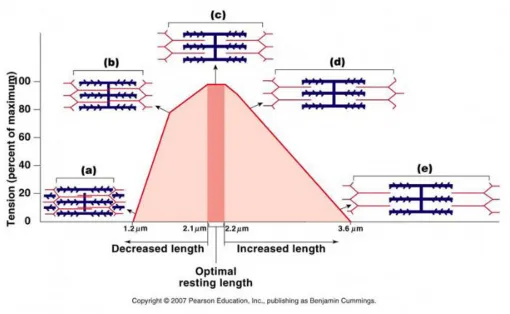
Figura da Internet: http://www.molecularlab.it: Relazione Tensione/Lunghezza.
Le fibre muscolari sviluppano forza in funzione della loro lunghezza iniziale e il sarcomero, unità funzionale del muscolo, si presta a rappresentare bene questo concetto. Esiste un ambito di lunghezze iniziali del sarcomero che permette di produrre la massima tensione. Al di sotto e al di sopra di questo ambito, la tensione sviluppata è inferiore e tende verso zero. Ciò si spiega con la probabilità di formazione dei ponti trasversi acto-miosinici; una sovrapposizione eccessiva fra i filamenti (lunghezze inferiori all’ottimale) o filamenti troppo sgranati (lunghezze superiori all’ottimale) intercettano valori di tensione più bassi perché diminuisce il numero dei ponti trasversi efficaci.
Tipologie di contrazione
La contrazione del muscolo striato è stata studiata da molto tempo, in vivo, in vitro, con modelli matematici, sull’uomo e su animali da esperimento, dagli aspetti più macroscopici sul campo a quelli a livello dei sarcomeri in laboratorio.
Sostanzialmente si fa riferimento a due tipi di contrazione che hanno luogo in vivo e possono essere ricreate in vitro: la contrazione isotonica e la contrazione isometrica.
Contrazione isotonica
Questa modalità indica un carico costante; il muscolo può dunque accorciarsi, cioè variare la propria lunghezza, mentre sostiene un carico. Come si è detto, in natura non esiste un carico nullo, mentre in laboratorio si può generare una contrazione isotonica a carico zero. Il muscolo si accorcia dunque, ad esempio, per flettere un avambraccio: il carico è rappresentato dal peso del segmento corporeo, che, ovviamente non varia, e questo gesto può essere compiuto a una determinata velocità, che è la massima consentita. Man mano che io sostengo con la mano pesi crescenti, la velocità di accorciamento è destinata a diminuire e il muscolo deve prima sostenere una fase in cui pareggiare la forza peso, sviluppando una tensione che chiameremo isometrica. Deve cioè prima raggiungere quella tensione a lunghezza costante e poi, con l’energia contrattile residua, spostare il segmento corporeo appesantito. Aumentando il carico si può arrivare a un peso talmente alto che, pur facendo sviluppare la massima forza al muscolo, non può più essere spostato. Con questo tipo di contrazione a carico crescente ci siamo spostati da una contrazione del tutto isotonica a una contrazione del tutto isometrica.
Contrazione isometrica
Questa modalità indica una lunghezza costante; il muscolo sviluppa forza in funzione del carico fino alla sua massima tensione isometrica. La velocità di accorciamento è nulla e il muscolo rimane alla lunghezza iniziale. Come si concilia questa tipologia di contrazione con la teoria dello scorrimento dei filamenti? Se il muscolo non varia la propria lunghezza non dovrebbero variarla nemmeno i sarcomeri, ma allora, come si può sviluppare forza? La risposta è che è la lunghezza esterna del muscolo a non cambiare, mentre all’interno i sarcomeri più centrali si accorciano a spese di quelli più distali, che vengono stirati, trasmettendo la forza ai capi del muscolo e permettendogli di sostenere un carico anche elevato.
Mentre la contrazione isotonica delle grandi masse muscolari (ad esempio gli arti inferiori) svolge il lavoro di pompa muscolare, aiuta cioè il ritorno venoso verso il cuore, consentendo così il mantenimento di una gittata cardiaca proporzionale all’esercizio dinamico, la contrazione isometrica opera una spremitura a valle, ma contestualmente può bloccare l’afflusso arterioso a monte, o ridurlo considerevolmente, date le tensioni sviluppate. La contrazione isometrica (statica) prosegue fondando il fabbisogno sull’aumento dell’estrazione di ossigeno dall’emoglobina e può essere necessario l’intervento della mioglobina, quando l’emoglobina desossigenata aumenta molto.
Contrazione isocinetica
Una contrazione isocinetica è una contrazione isotonica, ma a velocità costante, il che significa anche a potenza costante. Per fare un esempio, la differenza fra isotonica e isocinetica e quella che esiste fra l'esecuzione di 60 piegamenti sulle braccia e l'esecuzione di 60 piegamenti sulle braccia in 1 minuto: nel primo caso non c’è limite di tempo e la potenza può variare, in diminuzione probabilmente; nel secondo il gesto deve essere scandito dal tempo e la potenza deve rimanere costante, il che è più faticoso.
Fasi dell’isotonia
Durante la fase di accorciamento isotonico i vettori forza e lunghezza variano consensualmente: ad esempio in una flessione dell’avambraccio sono rivolti tutti e due verso l’alto: la lunghezza diminuisce avvicinando i capi muscolari e la forza sostiene il carico; viceversa nella fase di allungamento, mentre la lunghezza torna verso il valore iniziale, la forza rimane costante per continuare a sostenere il carico. La prima fase prende il nome di fase concentrica; la seconda fase prende il nome di fase eccentrica. Quest’ultima, se ripetuta a lungo da grandi masse muscolari, può anche essere dannosa. Fasi eccentriche importanti si manifestano nei muscoli quadricipiti femorali, durante il cammino o la corsa in discesa, dove il quadricipite, muscolo antigravitario, deve sostenere la sua quota di peso corporeo anche in allungamento. Una ricerca di molti anni fa aveva dimostrato che la corsa in discesa fa diminuire la massima tensione isometrica dei quadricipiti; un prelievo bioptico mostrava microlesioni a livello del reticolo sarcoplasmatico con minor disponibilità di Ca2+ per la contrazione.
Meccanismi energetici muscolari
-
Ana...