Biological Sciences
Amoebozoa
Amoebozoa is a diverse group of single-celled organisms characterized by their amoeboid movement and lack of a fixed shape. They are known for their ability to change their shape and engulf food particles by phagocytosis. Amoebozoa includes various species of amoebas and slime molds, and they play important roles in nutrient cycling and as predators in aquatic and soil ecosystems.
Written by Perlego with AI-assistance
Related key terms
1 of 5
11 Key excerpts on "Amoebozoa"
 eBook - ePub
eBook - ePub- David A. Baum, Douglas J. Futuyma, Hopi E. Hoekstra, Richard E. Lenski, Allen J. Moore, Catherine L. Peichel, Dolph Schluter, Michael C. Whitlock, David A. Baum, Douglas J. Futuyma, Hopi E. Hoekstra, Richard E. Lenski, Allen J. Moore, Catherine L. Peichel, Dolph Schluter, Michael C. Whitlock(Authors)
- 2013(Publication Date)
- Princeton University Press(Publisher)
chapter II.14 for fungi).The Amoebozoa include many of the organisms typically called “amoebae,” including the star of high school biology Amoeba proteus, two clades of slime molds, and Entamoeba histolytica, the causative agent of amoebic dysentery; however, this clade does not contain all amoebae, since amoeboid lineages can be found in virtually every major clade of eukaryotes. Amoebozoa emerged out of molecular phylogenetic analyses and are generally recovered in multigene molecular analyses, but there is no defining synapomorphy for Amoebozoa. The largest clade within the Amoebozoa, the Tubulinea, does have a morphological synapomorphy: cylindrical pseudopodia in which the cytoplasm streams in just one direction (monoaxial streaming). Examples of Tubulinea include the lobose testate (shelled) amoebae whose fossilized remains have been so useful in reconstructing paleoclimate and our old friend Amoeba.The Excavata were originally defined on the basis of a morphological and ultrastructural character, an “excavated” ventral feeding groove. The clade is only sometimes recovered, and then only in molecular analyses with many genes and dense taxonomic sampling. The most familiar Excavata are the causative agents of parasitic diseases. These include Giardia, a major cause of diarrhea worldwide that exacerbates malnutrition in children in the developing world. Other parasitic excavates include trypanosomes that cause sleeping sickness and Trichomonas vaginalis, which causes the sexually transmitted disease trichomoniasis. Members of Excavata are heterotrophic with the exception of one lineage of euglenids (a close relative of the trypanosomes) that acquired photosynthetic ability by engulfing a green alga endosymbiont hundreds of millions of years ago (see section 5 ). Many taxa, and all the parasites, within the Excavata are anaerobic and have highly reduced mitochondria, either hydrogenosomes or mitosomes. Some of these parasites also have rapid rates of evolution that artificially pulled them to the base of early ribosomal DNA trees (see chapter II.2 eBook - PDF
eBook - PDFCells and Their Component Parts
Biochemistry, Physiology, Morphology
- Jean Brachet, Alfred E. Mirsky(Authors)
- 2014(Publication Date)
- Academic Press(Publisher)
This process generally results in locomotion if the cells so engaged are attached to some substratum. In its simplest form, the cytoplasmic streaming pattern of an ameboid cell consists of an axial stream continuously displaced through a tubular body. There are many variations on this theme, however, which are still referred to as ameboid movement. While some amebae are predom-inantly monopodial, others send out many pseudopodia and are either temporarily or permanently polypodial. The shapes of pseudopodia vary in the extremes from the stout, almost cylindrical lobopodia of the larger species to the fine, straight, filamentous filopodia or the branching, an-astomosing reticulopodia of the foraminifera. Ameboid movement occurs in some form throughout Class Sarcodina, not only in the Order Amoebaea, where it is most commonly studied (free-living and parasitic amebae and testacea such as Difflugia and Arcella), but in the Orders Foraminifera and Rhizomastigina (amebo-flagellates) as well. The other orders in this class exhibit locomotion and protoplasmic streaming, but their movement probably should not be called ameboid according to the above definition. In the radiolaria and heliozoa, for example, protoplasmic streaming takes place along rigid skeletal spines. The proteomyxida (e.g., Labyrinthida; see Watson and Raper, 1957) are not strictly ameboid; they appear to slide along a slime track by an unknown mechanism. The slime molds are classified by the protozoologists as the Order Mycetozoa and by the botanists as Myxo-mycetes. During part of their life cycle they exist as myxamebae, and later fuse to form a plasmodium (e.g., Physarum polycephalum). Despite the fact that slime molds form from the fusion of single amebae and even in the plasmodial stage somewhat resemble huge amebae, the movements and streaming of plasmodia have been studied almost ex-clusively by plant physiologists. eBook - ePub
eBook - ePub- Tracey Lamb(Author)
- 2012(Publication Date)
- Wiley-Blackwell(Publisher)
Many of these species are commensals or opportunistic pathogens that only develop into medically important parasites in immunocompromised individuals. These species belong to five supergroups which are listed in underlined text, along with their mode of transmission, tissue tropism and global distribution. Species which were formally considered protozoa but are now known to be fungi are not listed.This chapter serves as an introduction to the world of protozoa. We have confined our discussion to the three protozoan supergroups that contain major human pathogens – the Amoebozoa, Excavata and Harosa. The general anatomy and life cycles of these pathogens will be summarised, as will the use of more recent innovations in genomic analysis that are reshaping our understanding of their biology and evolution.2.2 AmoebozoaThis supergroup consists of thirteen clades of amoebae and amoeboid flagellates. The Amoebozoa contains free-living, commensal, and parasitic species. Free-living species inhabit both terrestrial and aquatic environments. Most move via directed cytoplasmic flow and extension pseudopodia, although a few groups utilise flagella during particular points in their life cycle. Organisms within this group also show a great deal of diversity in their organelle complement. Some species have branching mitochondria with irregular cristae, degenerate or no mitochondria, or harbour a variety of endosymbionts. Several members of this group are parasites. of which Entamoeba histolytica is the most important to human health.2.2.1 EntamoebidaeThis family is comprised of species that are commensals or parasites of the digestive systems of arthropods or vertebrates. Genera and species have traditionally been differentiated based on nuclear morphology. The most important species, Entamoeba histolytica eBook - PDF
eBook - PDF- Kwang Jeon(Author)
- 2012(Publication Date)
- Academic Press(Publisher)
He wondered how amoebae were able to attach to a slide and how they could get longer or shorter without visible fibers. These questions are still the subject of research 130 years later, but we are inclined to rule out Dujardin's speculation that attachment might be controlled by the will of the little beings. B. NOMENCLATURE (cf. Chap. 2) The definition of Chaos, the generic name first proposed by Linnaeus, as a state of utter confusion or a state of things in which change is supreme (Webster) also provides a fitting description of the nomenclature relating to amoebae. From a historical point of view, it should be mentioned that the disagreements between the early naturalists as to which amoeba was being described were largely due to difficulties of communication as well as the generally elusive characters of amoebae. Even individuals belonging to the same strain vary tremendously in shape, size, and cytoplasmic inclusions, unless they are cultured under rigidly standardized conditions. An amoeba is largely 4 I. JOAN LORCH what it eats, and the specimens collected and described by the early observers must have reflected the nature of the food organisms available in their particular habitat as much as any characteristics of their own species. It is therefore not surprising that naturalists, who were as widely separated geographically as, for instance, Pénard and Leidy, and who often based their diagnosis on very few specimens without regard to cell age or state of nutrition, were apt to disagree on the definition of a particular species. Moreover, in the age of the electron 2 mm Fig. 1. Redrawn from Rôsel von Rosenhof (1755). The little Proteus. Drawing A is said to be natural size. The scale is approximate and applies to the rest of the figures. eBook - PDF
eBook - PDF- M Levendowsky(Author)
- 2012(Publication Date)
- Academic Press(Publisher)
Characterizing the forms treated in this chapter as small amebas leads to an inexact definition of the group. These organisms are often referred to as soil amebas, another descriptive term that tends to mislead. While found in soil, they also occur in aqueous habitats, as tissue culture contaminants, and as pathogens or potential pathogens in animal organisms. The group is diverse not only in regard to kinds of amebas, but also in regard to the ecological niches occupied by its individual members. We deal here with organisms that are (1) readily recovered from soil environments, (2) pygmies compared to ameboid giants such as Amoeba and Pelomyxa, (3) capable of morphogenetic transformation into a dor-mant cyst stage in all cases and, in some cases, a flagellate stage, (4) free-living but able, in several species, to develop as lethal pathogens when the opportunity presents itself and, finally, (5) difficult to define taxonomically because of a paucity of morphological features. Problems involving their identification and classification have kept these forms in relative obscurity, except for studies on soil ecology and differ-entiation. Attention shifted dramatically to this group over the past decade when some of these organisms were recognized as human pathogens. The accompanying notoriety, along with their utility in biochemical and devel-opmental studies, have earned the small amebas a niche of their very own, in contrast to their having shared a chapter with Entamoeba in the first edition of this series (Balamuth and Thompson, 1955). Excluded from consideration in this chapter are the cellular and true slime molds. The former are dealt with elsewhere in this series, and the latter are separated from the small amebas by a complex life cycle and other distinguishing features (Gray and Alexopoulos, 1968; Olive, 1975). The parasitic amebas, Entamoeba and its allies, are not treated in this chapter, nor are the various small amebas from marine habitats. Available until 19 Oct |Learn more
Available until 19 Oct |Learn moreIntelligent Evolution
How Alfred Russel Wallace's World of Life Challenged Darwinism
- Michael A. Flannery, Alfred Russel Wallace(Authors)
- 2020(Publication Date)
- Erasmus Press(Publisher)
One of the lowest or most rudimentary forms of life is the Amoeba, a living cell, just visible to the unaided eye as a little speck of floating jelly. This creature, being one of the most common of living microscopic objects, will have been seen by most of my readers. At first, under a low microscopic power, it appears structureless, as it was for some time described to be, but with increasing power and perfection of the microscope it is found to consist of three parts—a central body of a nearly globular shape slightly darker and more granular in texture, the outer jelly-like mass, and a small more transparent globular portion, which looks like an air-bubble, and is seen to undergo a slow motion of contraction and expansion; this is termed the “contractile vacuole,” which, when it has reached its full size, perhaps a quarter or a fifth of the whole diameter, suddenly disappears, and after a little while reappears and gradually grows again to its maximum size. The shape of the Amoeba varies greatly. Sometimes it is globular and immovable, but most frequently it is very irregular with arm-like processes jutting out in various directions. By careful watching, these are seen to increase or diminish so as to change the whole shape in an hour or two. But more curious is its power of absorbing any particles of organic matter that come in contact with it by gradually enclosing them in its substance, where after a time they disappear. The Amoebæ are found in stagnant water full of organic matter, and if they are transferred to pure water they soon diminish in size, proving that they require food and can digest it. The “contractile vacuole” is believed to have the function of expelling the carbonic acid gas and other waste products of assimilation.This Amoeba is one of the simplest forms of the lowest branch of the animal kingdom, the one-celled animals or Protozoa; all other animals being classed as Metazoa, as they are entirely built up of separate cells, which in all the more complex forms are countless millions in number. Every part of our bodies, from blood to muscles and nerves, from bones to skin, hair, and nails, is alike constructed of variously modified cells.It might be thought that animals consisting of single cells could not be very numerous or very differently organised. Yet they are grouped into five classes, the first, Rhizopoda, comprising not only many kinds of Amoebae, but the beautiful Foraminifera, whose exquisite shells are such favourite microscopic objects. They are single amoeboid cells which yet have the power either of building up shells of small inorganic particles, or of secreting the more beautiful shells which seem to mimic the forms of those of the higher Mollusca. The fossils called Nummulites were Foraminifera with flat coiled shells, forming great masses of Eocene limestone. They are the largest of all, some equalling a half-crown in size. Radiolaria are rhizopods having a beautiful siliceous skeleton, and often living in colonies. Another class, the Mastigophora, have extremely varied shapes, often like sea-weed or flowers, having long, slender, whip-like processes. These and hundreds of other strange forms are still essentially single cells, though often grouped together for a time, and they all increase either by division or by giving off buds, which rapidly grow into the perfect form. eBook - ePub
eBook - ePubThorp and Covich's Freshwater Invertebrates
Keys to Nearctic Fauna
- James H. Thorp, D. Christopher Rogers(Authors)
- 2015(Publication Date)
- Academic Press(Publisher)
2.1 B). There is considerable diversity of structure in the amoebae, particularly in the character of any shell or skeletal material that may be present, and in the type of pseudopodium, for example, broadly lobed, needle-like, or reticulate. Amoebae range in size from only a few micrometers to 2 mm in diameter. Although many lack a fixed external morphology, the characteristic morphologies shown by the various taxa are surprisingly distinctive, even if difficult to quantify (Fig. 2.4). By using also the number, size, and structure of organelles and characteristics of tests (where present), identification is not as difficult for living specimens as might be imagined. The morphology of amoebae is plastic. Many adopt a stellate morphology if suspended in water, but few are truly planktonic; rather, they live on surfaces or in sediments. In most, for example, Amoeba (Figs. 2.1 B and 2.4 F), the cytoplasm is divided into an inner granular endoplasm and an outer hyaline ectoplasm, or hyaloplasm, with a characteristic thickness and distribution around the cell. Locomotion may be achieved by extending many pseudopodia simultaneously, as in Amoeba (Figs. 2.1 B and 2.4 F), or by moving as a single mass on a broad front (2.4 E, P), or as a cylinder (limax amoebae, Fig. 2.4 I, K, L). Not only do pseudopodia have characteristic shapes, but the tail end or uroid may be distinctive (Fig. 2.4 J, L), and the cell surface may be distinctly sculptured, as in Thecamoeba (Fig. 2.4 D). The classification of the naked, lobose amoebae was recently revised by Smirnov et al. (2011). Other groups of amoebae, notably the testate amoebae, possess shells (or tests) that may be proteinaceous, agglutinate, siliceous or calcareous in composition (Figs. 2.5 A–Q and 2.6 B–K). These are generally vase-shaped, with a single opening through which pseudopodia emerge. Many are terrestrial, but benthic forms are common, and a few are planktonic eBook - ePub
eBook - ePubAnimals Without Backbones
An Introduction to the Invertebrates
- Ralph Buchsbaum, Mildred Buchsbaum, John Pearse, Vicki Pearse(Authors)
- 2013(Publication Date)
- University of Chicago Press(Publisher)
Amoeba proteus. Photomicrograph. (R. Buchsbaum)A giant among amebas is Chaos carolinense, which can reach a length of 5 mm. This and related species are unusual in having numerous nuclei (large white dots among smaller granules) and many contractile vacuoles, presumably associated with large size. Although one-celled, this large ameba dwarfs the many-celled rotifer (see chapter 14 ) beside it. Photographed under dark-field illumination, the cell membrane and cytoplasm appear white on a black background. Photomicrograph. (P. S. Tice)Most protozoans consist of a single cell, organized much like one cell in a many-celled animal. Yet a protozoan behaves much as a whole animal does and carries out essentially the same life activities. The internal organization of some protozoans is more complex than that of any single cell in a many-celled animal. Such protozoans have elaborate mechanisms for moving, feeding, defending themselves, excreting, coordinating activities, and performing sexual reproduction—all within the limits of a single cell. We shall examine one of these later in this chapter. We will begin, however, with some simpler protozoans that are perhaps more surprising, in that they seem to have so little permanent structure or differentiated organization. Yet they manage to go on about their activities without any of the fancy equipment that most other organisms require to survive.A Simple Protozoan: AmebaThe typical ameba of freshwater ponds is usually of microscopic size, but large individuals may reach half a millimeter in diameter, being visible to the naked eye as white specks. Each ameba is a little mass of clear gelatinous material containing many granules and droplets. The surface is bounded by a delicate cell membrane through which materials pass in and out of the ameba. The membrane possesses the important property of selective permeability,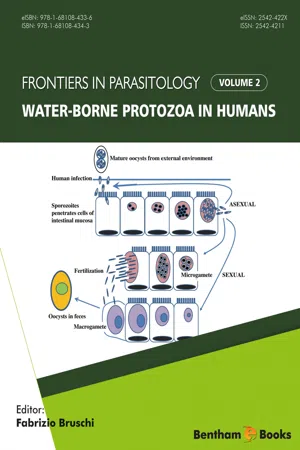 eBook - ePub
eBook - ePub- Fabrizio Bruschi(Author)
- 2017(Publication Date)
- Bentham Science Publishers(Publisher)
Free-living AmoebaeFree-living amoebae (FLA) are aerobic, mitochondriate, eukaryotic protists and have the capability to exist as free- living organisms in nature and invading living host so they are also called as amphizoic amoebae [1 ]. The present classification schemes of protists, based largely on their genetic relatedness while the traditional classification scheme which is based on morphological characteristics, is no longer a valid scheme. According to the new classification scheme, the Eukaryotes have been classified into six clusters or ‘Super Groups’, namely Amoebozoa, Opisthokonta, Rhizaria, Archaeplastida, Chromalveolata and Excavata. The four amoebaehave been classified into two Super Groups, Amoebozoa and Excavata, Acanthamoeba and Balamuthia are under cluster Amoebozoa: Acanthamoebidae; while Naegleria fowleri comes under excavata: Heterolobosia: Vahlkampfiidae; and Sappinia under the cluster Amoebozoa: Flabellinea: Thecamoebidae [2 ]. There is an increasing scientific interest in the field of free-living amoebae as determined by published articles over the last five decade. A pubmed search using “Acanthamoeba”, “Balamuthia”, Naegleria” or “Sappinia” was carried out from 1960 to 2010 which showed constant increase in number of cases from zero to nearly two hundred. Now, the question arises, why they are on the rise? There are many explanations given. Number of immunocompromised patients and contact lens wearers are on the rise [2 ]. Ability to act as a host or reservoir for other microbial pathogens [3 ]. These are model organism for motility studies which has led to a significant interest in this organism over the years. Ecologically, infections with free living amoebae have been linked to warm water such as above-ground pipelines, tropical lakes, geothermal water, heated swimming pools or discharges of industrial cooling water and soil [2 ]. Free living amoeba plays two major ecological roles: Influencing the structure of the microbial community and enhancing nutrient recycling. Both these activities are associated with free living amoebae (FLA). They feed on bacteria thus regulating bacterial populations in the soil. These FLAs cause 60% of the total reduction in bacterial population. Bacteria are primary decomposers of organic material but fail to release minerals from their mass, FLAs act as secondary decomposers thus releasing minerals that are tied up with bacteria. In this way, FLAs make the nutrients available that would otherwise remain inaccessible. The soil containing Acanthamoeba and bacteria showed significantly greater mineralization of carbon, nitrogen, and phosphorous compared to the soil containing bacteria without FLAs. Overall, these FLAs regulate the population of bacteria in ecosystem and this was demonstrated by the findings that nitrogen mineralization by Acanthamoeba permitted continued growth of bacteria such as Pseudomonas paucimobilis resulting in a greater bacterial biomass [3 , 4 eBook - PDF
eBook - PDF- J. K. Misra, Jalpa P. Tewari, Sunil Kumar Deshmukh, Csaba Vágvölgyi, J. K. Misra, Jalpa P. Tewari, Sunil Kumar Deshmukh, Csaba Vágvölgyi(Authors)
- 2014(Publication Date)
- CRC Press(Publisher)
Their phylogenetic position is currently supported by molecular studies that clearly show their monophyletic character within the Amoebozoa (see Pawlowski and Burki, 2009). In the past, myxomycetes usually have been considered as members of a group known as the Eumycetozoa (Olive, 1975). This hypothetical group included, along with the myxomycetes, two other groups of amoebae known as dictyostelids and protostelids (also called protosteloid amoebae). However, the integrity of the Eumycetozoa as a natural group has been questioned recently on the basis of molecular evidence indicating that the protostelids are probably not monophyletic (Shadwick et al., 2009). This fact does not affect the currently recognized position of myxomycetes within the Amebozoa, but it changes the concepts and nomenclatural treatments of the particular subgroups of organisms to which they are related. Given this situation and for the purpose of this chapter, myxomycetes simply will be treated herein as a group of Amoebozoans taxonomically distinct from both dictyostelids and protostelids. Life Cycle The generalized (“textbook”) life cycle of myxomycetes (Fig. 6.1) has been described in detail by a number of authors (e.g., Martin and Alexopoulos, 1969; Stephenson and Stempen, 1994; Everhart and Keller, 2008). In brief, the life cycle involves two trophic stages—one consisting of microscopic uninucleate amoeboflagellate cells and the other of a multinucleate plasmodium—along with a reproductive stage (fruiting body or sporocarp) somewhat similar to the spore-producing fruiting bodies of certain macrofungi, albeit much smaller. Although myxomycetes are fundamentally amoeboid organisms, their capability to produce a fruiting body that is large enough to be observed directly in nature (the exceptionally large fruiting bodies produced by some species can be the size of a dinner plate!) is truly remarkable (Martin and Alexopoulos, 1969; Stephenson and Stempen, 1994). eBook - PDF
eBook - PDF- Mahmut Caliskan(Author)
- 2012(Publication Date)
- IntechOpen(Publisher)
5. Taxonomy of Heterolobosea The taxon Heterolobosea was created by Page & Blanton (1985) as a class unifying orders Schizopyrenida (limax-type amoebae, often with the flagellate stage) and Acrasida (aggregative amoebae forming multicellular sorocarps) on the basis of the common presence Genetic Diversity in Microorganisms 12 of limax amoeba with eruptive lobopodia, discoidal mitochondrial cristae, and the absence of a stacked Golgi apparatus. However, it was later shown that several organisms with different morphology are closely related to the Heterolobosea ( Pharyngomonas ) or even form its internal branches ( Percolomonas , Stephanopogon , Lyromonas , Psalteriomonas , Pleurostomum ). Currently, two concepts of Heterolobosea, here called Heterolobosea sensu lato and Heterolobosea sensu stricto , respectively, exist. The concept of Heterolobosea sensu lato emphasizes monophyletic taxa and includes all aforementioned genera in Heterolobosea. It means, in fact, that Heterolobosea sensu lato is a group containing all descendants of the last common ancestor of Pharyngomonas and Naegleria . This concept is currently favored by most authors and we follow it as well. In contrast, some authors emphasize the original definition of Heterolobosea sensu Page & Blanton (1985). The absence of mitochondrial cristae and microbodies in Lyromonas , Sawyeria , and Monopylocystis , the presumed absence of the amoeboid stage in Percolomonas , Stephanopogon and Pharyngomonas (but not in Pleurostomum ), and the different arrangement of flagella of Pharyngomonas are considered so important that these genera cannot be members of Heterolobosea. Instead, they are classified in separate classes closely related to the Heterolobosea sensu stricto . The taxon corresponding to the Heterolobosea sensu lato was named Percolozoa (see Cavalier-Smith, 1991, 1993, 2003). The most recent version of this concept is represented by Cavalier-Smith & Nikolaev (2008).
Index pages curate the most relevant extracts from our library of academic textbooks. They’ve been created using an in-house natural language model (NLM), each adding context and meaning to key research topics.